
Recent Studies
Formation Principles (together with Lia Addadi)
In the almost 25 years since the late Heinz Lowenstam and I published the book entitled “On Biomineralization” (Lowenstam and Weiner, 1989) some fundamental concepts still prevail (eg the preformed extracellular matrix into which mineral forms), but some basic paradigms have changed. The concept that the first minerals are often deposited in vesicles within cells and are then transported into the extracellular space where they often undergo a transformation into more ordered forms, is rapidly becoming established (Fig. 1). This idea goes back to the work of Towe and Lowenstam (Towe and Lowenstam, 1967), but gained tremendous momentum when Beniash et al ((Beniash et al., 1997) demonstrated that sea urchin larval spicules form via an amorphous calcium carbonate precursor phase that is produced intracellularly. In recent years Beniash et al (Beniash et al., 2009) demonstrated that this is also the strategy used in the formation of vertebrate enamel, and Mahamid et al (Mahamid et al., 2010) showed that vertebrate bone also forms via an amorphous calcium phosphate (or something similar) phase. This phase initially forms in cells and is then transported out to the extracellular environment. There are also interesting variations on this theme. Certain Foraminiferidae and Coccolithophoridae form the mature mineralized product inside intracellular vesicles and only then extrude them into the extracellular environment (Lowenstam and Weiner, 1989), and some organisms use the mature product while still in the cell. A fascinating example of the latter is the widespread production of guanine crystals inside vesicles and the arrangement of the crystal bearing vesicles in stacks in order to produce impressive photonic colors. Fish skin (Levy-Lior et al., 2007), spider cuticles (Levy-Lior et al., 2010) and even some amazing copepods (Chae et al., 1996) utilize this strategy. Biogenic guanine also forms via an amorphous precursor phase (Gur et al., 2012). For an overview, read the review on biomineralization pathways (Weiner and Addadi, 2011).
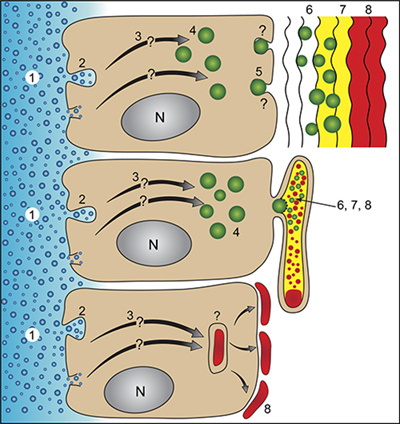 |
Figure 1: Schemes of generalized crystallization pathways involving an extracellular matrix (top), a vesicle confined space (syncitium) (middle), and the formation of mature mineralized elements within a vesicle inside the cell (bottom). The numbers designate the following stages in the crystallization pathways: 1. The medium from which the ions are derived (sea water or body fluids). 2. The ion sequestering process – endocytosis of sea water droplets and/or ion channels and/or transporters. 3. Transport within the cell to specialized vesicles. 4. Specialized vesicles in which the formation of the first disordered mineral phase occurs. 5. Transport of the mineral-bearing vesicles and their contents into the extracellular environment or into the syncitium. 6. The translocation of the disordered phase to the crystallization front. 7. Transformation of the initial disordered phase into more ordered phases. 8. The mature mineralized tissue. In the case of the bottom crystallization pathway, the mature mineralized product may remain within the cell (eg.guanine crystals in fish skin) or may be transported to the cell surface (Coccolithophoridae and miliolid foraminifera). Very little is known about transient precursor phases in the bottom crystallization pathway. (From Weiner and Addadi, 2011) |
The widespread utilization by organisms of a disordered precursor phase, has sparked much renewed interest in this well known chemical phenomenon espoused initially by Ostwald (Ostwald, 1897). Perhaps the most significant outcome is the discovery by Colfen and colleagues that ion clusters can exist in a stable state even in solutions that are undersaturated (Gebauer et al., 2008). There is however much still to understand regarding this precursor phase: how the ions are transported into the cells, the precise mineral form in which they are initially formed, and how once in the extracellular space they crystallize. One key instrument for understanding aspects of this complex process, is Xray Photon Emmision Electron Microscopy (XPEEM). XPEEM was used to monitor the transformation from ACC to calcite in the sea urchin larva (Politi et al., 2008) (Fig. 2).
 |
Figure 2: Schematic illustration of a hypothetical pathway (yellow arrows) for the propagation of crystallization in a sea urchin spicule by secondary nucleation among ACC particles. The substrate is an etched fractured section of a spine showing the profiles of the original particles, the blue triangle denotes the hypothetical original calcite crystal, and the red circles denote particles that remain excluded from the crystallization path. (From Weiner and Addadi, 2011) |