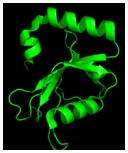
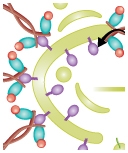
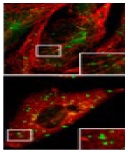
Understanding the process of intracellular protein transport, whereby protein-carrying vesicles move across and between membranes, is of fundamental importance in cell physiology and medicine. Autophagy (literally self-eating) is a unique membrane trafficking process essential for the degradation and recycling of excess and/or defective macromolecules and organelles in eukaryotes. This pathway is activated under environmental stress conditions as well as during certain developmental stages and has been linked to programmed cell death, cancer, pathogen infection, and degradation of ubiquitinated protein aggregates formed in many pathological conditions. The main goal of our studies is to understand the molecular mechanism of autophagy and characterize its exact role in health and disease employing mammalian, and yeast systems.