Alber S., Di Matteo P., Zdradzinski M. D., Costa I. D., Medzihradszky K. F., Kawaguchi R., Di Pizio A., Freund P., Panayotis N., Marvaldi L., Doron-Mandel E., Okladnikov N., Rishal I., Nevo R., Coppola G., Joon Lee S., Sahoo P. K., Burlingame A. L., Twiss J. L. & Fainzilber M.
(2023)
Science advances.
9,
30,
adi0286.
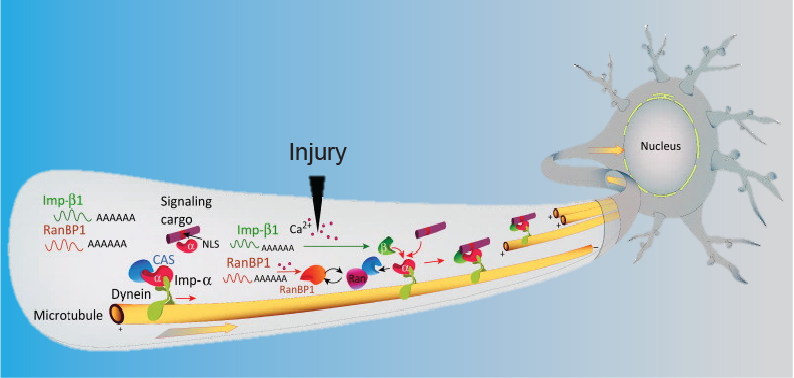
Polypyrimidine tract binding protein 1 (PTBP1) is thought to be expressed only at embryonic stages in central neurons. Its down-regulation triggers neuronal differentiation in precursor and non-neuronal cells, an approach recently tested for generation of neurons de novo for amelioration of neurodegenerative disorders. Moreover, PTBP1 is replaced by its paralog PTBP2 in mature central neurons. Unexpectedly, we found that both proteins are coexpressed in adult sensory and motor neurons, with PTBP2 restricted mainly to the nucleus, while PTBP1 also shows axonal localization. Levels of axonal PTBP1 increased markedly after peripheral nerve injury, and it associates in axons with mRNAs involved in injury responses and nerve regeneration, including importin β1 (KPNB1) and RHOA. Perturbation of PTBP1 affects local translation in axons, nociceptor neuron regeneration and both thermal and mechanical sensation. Thus, PTBP1 has functional roles in adult axons. Hence, caution is required before considering targeting of PTBP1 for therapeutic purposes.
Doron E., Koppel I., Abraham O., Rishal I., Smith T. P., Buchanan C. N., Sahoo P. K., Kadlec J., Oses-Prieto J. A., Kawaguchi R., Alber S., Zahavi E. E., Di Matteo P., Di Pizio A., Song D. A., Okladnikova N., Gordon D., Ben-Dor S., Haffner-Krausz R., Coppola G., Burlingame A. L., Jungwirth P., Twiss J. L. & Fainzilber M.
(2021)
EMBO Journal.
40,
20,
e107158.
Nucleolin is a multifunctional RNA Binding Protein (RBP) with diverse subcellular localizations, including the nucleolus in all eukaryotic cells, the plasma membrane in tumor cells, and the axon in neurons. Here we show that the glycine arginine rich (GAR) domain of nucleolin drives subcellular localization via protein-protein interactions with a kinesin light chain. In addition, GAR sequences mediate plasma membrane interactions of nucleolin. Both these modalities are in addition to the already reported involvement of the GAR domain in liquid-liquid phase separation in the nucleolus. Nucleolin transport to axons requires the GAR domain, and heterozygous GAR deletion mice reveal reduced axonal localization of nucleolin cargo mRNAs and enhanced sensory neuron growth. Thus, the GAR domain governs axonal transport of a growth controlling RNA-RBP complex in neurons, and is a versatile localization determinant for different subcellular compartments. Localization determination by GAR domains may explain why GAR mutants in diverse RBPs are associated with neurodegenerative disease.
Terenzio M., Koley S., Samra N., Rishal I., Zhao Q., Sahoo P. K., Urisman A., Marvaldi L., Oses-Prieto J. A., Forester C., Gomes C., Kalinski A. L., Di Pizio A., Doron-Mandel E., Perry R. B. T., Koppel I., Twiss J. L., Burlingame A. L. & Fainzilber M.
(2018)
Science.
359,
6382,
p. 1416-1421
How is protein synthesis initiated locally in neurons? We found that mTOR (mechanistic target of rapamycin) was activated and then up-regulated in injured axons, owing to local translation of mTOR messenger RNA (mRNA). This mRNA was transported into axons by the cell size-regulating RNA-binding protein nucleolin. Furthermore, mTOR controlled local translation in injured axons. This included regulation of its own translation and that of retrograde injury signaling molecules such as importin b1 and STAT3 (signal transducer and activator of transcription 3). Deletion of the mTOR 3 untranslated region (3UTR) in mice reduced mTOR in axons and decreased local translation after nerve injury. Both pharmacological inhibition of mTOR in axons and deletion of the mTOR 3UTR decreased proprioceptive neuronal survival after nerve injury. Thus, mRNA localization enables spatiotemporal control of mTOR pathways regulating local translation and long-range intracellular signaling.
Perry R. B. T., Rishal I., Doron-Mandel E., Kalinski A. L., Medzihradszky K. F., Terenzio M., Alber S., Koley S., Lin A., Rozenbaum M., Yudin D., Sahoo P. K., Gomes C., Shinder V., Geraisy W., Huebner E. A., Woolf C. J., Yaron A., Burlingame A. L., Twiss J. L. & Fainzilber M.
(2016)
Cell Reports.
16,
6,
p. 1664-1676
How can cells sense their own size to coordinate biosynthesis and metabolism with their growth needs? We recently proposed a motor-dependent bidirectional transport mechanism for axon length and cell size sensing, but the nature of the motor-transported size signals remained elusive. Here, we show that motor-dependent mRNA localization regulates neuronal growth and cycling cell size. We found that the RNA-binding protein nucleolin is associated with importin β1 mRNA in axons. Perturbation of nucleolin association with kinesins reduces its levels in axons, with a concomitant reduction in axonal importin β1 mRNA and protein levels. Strikingly, subcellular sequestration of nucleolin or importin β1 enhances axonal growth and causes a subcellular shift in protein synthesis. Similar findings were obtained in fibroblasts. Thus, subcellular mRNA localization regulates size and growth in both neurons and cycling cells.
Rishal I., Kam N., Perry R. B., Shinder V., Fisher E. M. C., Schiavo G. & Fainzilber M.
(2012)
Cell Reports.
1,
6,
p. 608-616
Size homeostasis is fundamental in cell biology, but it is not clear how large cells such as neurons can assess their own size or length. We examined a role for molecular motors in intracellular length sensing. Computational simulations suggest that spatial information can be encoded by the frequency of an oscillating retrograde signal arising from a composite negative feedback loop between bidirectional motor-dependent signals. The model predicts that decreasing either or both anterograde or retrograde signals should increase cell length, and this prediction was confirmed upon application of siRNAs for specific kinesin and/or dynein heavy chains in adult sensory neurons. Heterozygous dynein heavy chain 1 mutant sensory neurons also exhibited increased lengths both in vitro and during embryonic development. Moreover, similar length increases were observed in mouse embryonic fibroblasts upon partial downregulation of dynein heavy chain 1. Thus, molecular motors critically influence cell-length sensing and growth control.