Keret S., Meltzer H., Marmor-Kollet N. & Schuldiner O.
(2025)
Life Science Alliance.
8,
12,
e202503504.
Schizophrenia is a multifactorial neuropsychiatric disorder of complex and mostly unknown etiology, affected by genetic, developmental, and environmental factors. Neuroanatomical abnormalities, such as loss of gray matter, are apparent before the onset of symptoms, suggesting a neurodevelopmental origin. Indeed, it has been hypothesized, and recently experimentally supported, that schizophrenia is associated with dysregulation of developmental synaptic pruning. Here, we explore the molecular link between schizophrenia-associated genes and developmental neuronal remodeling. We focused on the Drosophila mushroom body, which undergoes stereotypic remodeling during metamorphosis. We conducted a loss-of-function screen in either glia or neurons of Drosophila homologs of human genes that are associated with schizophrenia based on genomic studies. Out of our \u201cpositive hits,\u201d we focused on matrix metalloproteinases. Our combinatorial loss-of-function experiments suggest that Drosophila matrix metalloproteinases are required both in neurons and in glia for the pruning of mushroom body axons. Our results shed new light on potential molecular players underlying neurodevelopmental defects in schizophrenia and highlight the advantage of genetically tractable model organisms in the study of human neurodevelopmental disorders.
Morano N. C., Lopez D. H., Meltzer H., Sergeeva A. P., Katsamba P. S., Rostam K. D., Gupta H. P., Becker J. E., Bornstein B., Cosmanescu F., Schuldiner O., Honig B., Mann R. S. & Shapiro L.
(2025)
PLoS Biology.
23,
3,
e3003030.
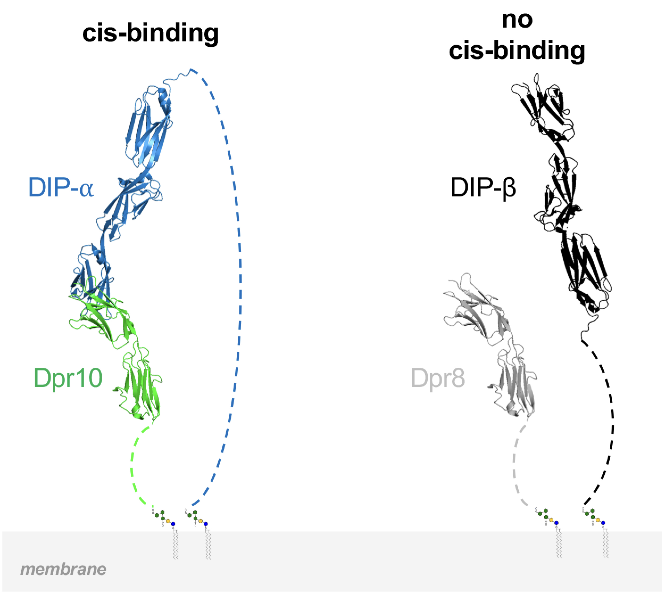
In Drosophila, two interacting adhesion protein families, Defective proboscis responses (Dprs) and Dpr interacting proteins (DIPs), coordinate the assembly of neural networks. While intercellular DIP::Dpr interactions have been well characterized, DIPs and Dprs are often co-expressed within the same cells, raising the question as to whether they also interact in cis. We show, in cultured cells and in vivo, that DIP-α and DIP-δ can interact in cis with their ligands, Dpr6/10 and Dpr12, respectively. When co-expressed in cis with their cognate partners, these Dprs regulate the extent of trans binding, presumably through competitive cis interactions. We demonstrate the neurodevelopmental effects of cis inhibition in fly motor neurons and in the mushroom body. We further show that a long disordered region of DIP-α at the C-terminus is required for cis but not trans interactions, likely because it alleviates geometric constraints on cis binding. Thus, the balance between cis and trans interactions plays a role in controlling neural development.
Çoban B., Poppinga H., Rachad E. Y., Geurten B., Vasmer D., Jimenez F. J. R., Gadgil Y., Deimel S. H., Alyagor I., Schuldiner O., Grunwald Kadow I. C., Riemensperger T. D., Widmann A. & Fiala A.
(2024)
Learning and Memory.
31,
5,
p. 1-15
Associative learning enables the adaptive adjustment of behavioral decisions based on acquired, predicted outcomes. The valence of what is learned is influenced not only by the learned stimuli and their temporal relations, but also by prior experiences and internal states. In this study, we used the fruit fly Drosophila melanogaster to demonstrate that neuronal circuits involved in associative olfactory learning undergo restructuring during extended periods of low-caloric food intake. Specifically, we observed a decrease in the connections between specific dopaminergic neurons (DANs) and Kenyon cells at distinct compartments of the mushroom body. This structural synaptic plasticity was contingent upon the presence of allatostatin A receptors in specific DANs and could be mimicked optogenetically by expressing a light-activated adenylate cyclase in exactly these DANs. Importantly, we found that this rearrangement in synaptic connections influenced aversive, punishment-induced olfactory learning but did not impact appetitive, reward-based learning. Whether induced by prolonged low-caloric conditions or optogenetic manipulation of cAMP levels, this synaptic rearrangement resulted in a reduction of aversive associative learning. Consequently, the balance between positive and negative reinforcing signals shifted, diminishing the ability to learn to avoid odor cues signaling negative outcomes. These results exemplify how a neuronal circuit required for learning and memory undergoes structural plasticity dependent on prior experiences of the nutritional value of food.
Mayseless O., Shapira G., Rachad E. Y., Fiala A. & Schuldiner O.
(2023)
Current Biology.
33,
5,
p. 981-989.e3
Postnatal remodeling of neuronal connectivity shapes mature nervous systems.1,2,3 The pruning of exuberant connections involves cell-autonomous and non-cell-autonomous mechanisms, such as neuronal activity. Indeed, experience-dependent competition sculpts various excitatory neuronal circuits.4,5,6,7,8,9 Moreover, activity has been shown to regulate growth cone motility and the stability of neurites and synaptic connections.10,11,12,13,14 However, whether inhibitory activity influences the remodeling of neuronal connectivity or how activity influences remodeling in systems in which competition is not clearly apparent is not fully understood. Here, we use the Drosophila mushroom body (MB) as a model to examine the role of neuronal activity in the developmental axon pruning of γ-Kenyon cells. The MB is a neuronal structure in insects, implicated in associative learning and memory,15,16 which receives mostly olfactory input from the antennal lobe.17,18 The MB circuit includes intrinsic neurons, called Kenyon cells (KCs), which receive inhibitory input from the GABAergic anterior paired lateral (APL) neuron among other inputs. The γ-KCs undergo stereotypic, steroid-hormone-dependent remodeling19,20 that involves the pruning of larval neurites followed by regrowth to form adult connections.21 We demonstrate that silencing neuronal activity is required for γ-KC pruning. Furthermore, we show that this is mechanistically achieved by cell-autonomous expression of the inward rectifying potassium channel 1 (irk1) combined with inhibition by APL neuron activity likely via GABA-B-R1 signaling. These results support the Hebbian-like rule \u201cuse it or lose it,\u201d where inhibition can destabilize connectivity and promote pruning while excitability stabilizes existing connections.
Marmor-Kollet N., Berkun V., Cummings G., Keren-Shaul H., David E., Addadi Y. & Schuldiner O.
(2023)
Cell Reports.
42,
2,
112117.
Astrocytes are essential for synapse formation, maturation, and plasticity; however, their function during developmental neuronal remodeling is largely unknown. To identify astrocytic molecules required for axon pruning of mushroom body (MB) γ neurons in Drosophila, we profiled astrocytes before (larva) and after (adult) remodeling. Focusing on genes enriched in larval astrocytes, we identified 12 astrocytic genes that are required for axon pruning, including the F-actin regulators Actin-related protein 2/3 complex, subunit 1 (Arpc1) and formin3 (form3). Interestingly, perturbing astrocytic actin dynamics does not affect their gross morphology, migration, or transforming growth factor β (TGF-β) secretion. In contrast, actin dynamics is required for astrocyte infiltration into the axon bundle at the onset of pruning. Remarkably, decreasing axonal adhesion facilitates infiltration by Arpc1 knockdown (KD) astrocytes and promotes axon pruning. Conversely, increased axonal adhesion reduces lobe infiltration by wild-type (WT) astrocytes. Together, our findings suggest that actin-dependent astrocytic infiltration is a key step in axon pruning, thus promoting our understanding of neuron-glia interactions during remodeling.
Poppinga H., Coban B., Meltzer H., Mayseless O., Widmann A., Schuldiner O. & Fiala A.
(2022)
Open Biology.
12,
9,
220096.
The principles of how brain circuits establish themselves during development are largely conserved across animal species. Connections made during embryonic development that are appropriate for an early life stage are frequently remodelled later in ontogeny via pruning and subsequent regrowth to generate adult-specific connectivity. The mushroom body of the fruit fly Drosophila melanogaster is a well-established model circuit for examining the cellular mechanisms underlying neurite remodelling. This central brain circuit integrates sensory information with learned and innate valences to adaptively instruct behavioural decisions. Thereby, the mushroom body organizes adaptive behaviour, such as associative learning. However, little is known about the specific aspects of behaviour that require mushroom body remodelling. Here, we used genetic interventions to prevent the intrinsic neurons of the larval mushroom body (gamma-type Kenyon cells) from remodelling. We asked to what degree remodelling deficits resulted in impaired behaviour. We found that deficits caused hyperactivity and mild impairment in differential aversive olfactory learning, but not appetitive learning. Maintenance of circadian rhythm and sleep were not affected. We conclude that neurite pruning and regrowth of gamma-type Kenyon cells is not required for the establishment of circuits that mediate associative odour learning per se, but it does improve distinct learning tasks.
Meltzer H. & Schuldiner O.
(2022)
Frontiers in Neuroscience.
16,
897706.
Developmental neuronal remodeling is required for shaping the precise connectivity of the mature nervous system. Remodeling involves pruning of exuberant neural connections, often followed by regrowth of adult-specific ones, as a strategy to refine neural circuits. Errors in remodeling are associated with neurodevelopmental disorders such as schizophrenia and autism. Despite its fundamental nature, our understanding of the mechanisms governing neuronal remodeling is far from complete. Specifically, how precise spatiotemporal control of remodeling and rewiring is achieved is largely unknown. In recent years, cell adhesion molecules (CAMs), and other cell surface and secreted proteins of various families, have been implicated in processes of neurite pruning and wiring specificity during circuit reassembly. Here, we review some of the known as well as speculated roles of CAMs in these processes, highlighting recent advances in uncovering spatiotemporal aspects of regulation. Our focus is on the fruit fly Drosophila, which is emerging as a powerful model in the field, due to the extensive, well-characterized and stereotypic remodeling events occurring throughout its nervous system during metamorphosis, combined with the wide and constantly growing toolkit to identify CAM binding and resulting cellular interactions in vivo. We believe that its many advantages pose Drosophila as a leading candidate for future breakthroughs in the field of neuronal remodeling in general, and spatiotemporal control by CAMs specifically.
Bornstein B., Meltzer H., Adler R., Alyagor I., Berkun V., Cummings G., Reh F., Keren-Shaul H., David E., Riemensperger T. & Schuldiner O.
(2021)
EMBO Journal.
40,
12,
e105763.
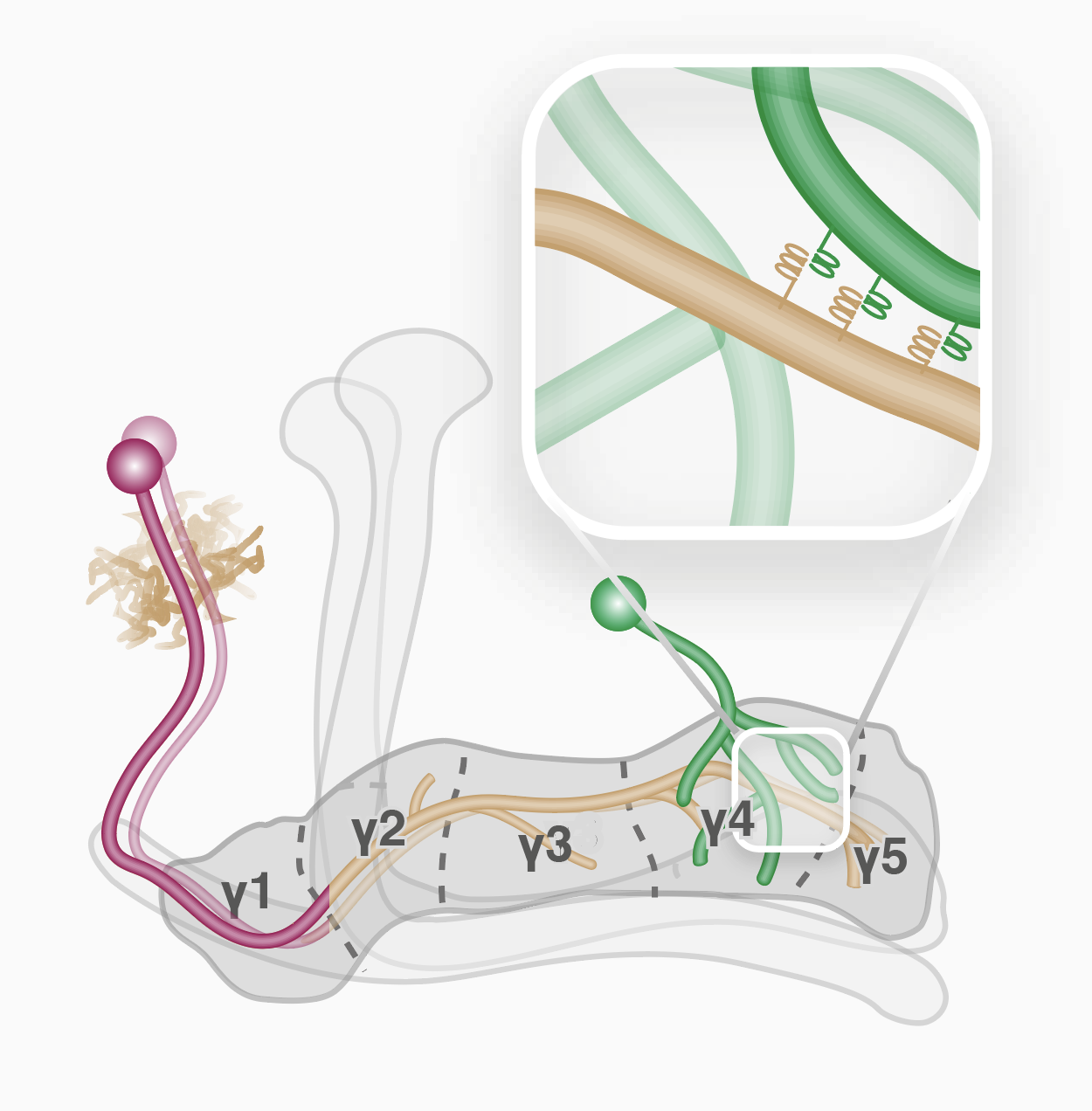
The mechanisms controlling wiring of neuronal networks are not completely understood. The stereotypic architecture of the Drosophila mushroom body (MB) offers a unique system to study circuit assembly. The adult medial MB γ-lobe is comprised of a long bundle of axons that wire with specific modulatory and output neurons in a tiled manner, defining five distinct zones. We found that the immunoglobulin superfamily protein Dpr12 is cell-autonomously required in γ-neurons for their developmental regrowth into the distal γ4/5 zones, where both Dpr12 and its interacting protein, DIP-δ, are enriched. DIP-δ functions in a subset of dopaminergic neurons that wire with γ-neurons within the γ4/5 zone. During metamorphosis, these dopaminergic projections arrive to the γ4/5 zone prior to γ-axons, suggesting that γ-axons extend through a prepatterned region. Thus, Dpr12/DIP-δ transneuronal interaction is required for γ4/5 zone formation. Our study sheds light onto molecular and cellular mechanisms underlying circuit formation within subcellular resolution.
Meltzer H. & Schuldiner O.
(2020)
Current Opinion in Neurobiology.
63,
p. 23-30
Developmental neuronal remodeling shapes the mature connectivity of the nervous system in both vertebrates and invertebrates. Remodeling often combines degenerative and regenerative events, and defects in its normal progression have been linked to neurological disorders. Here we review recent progress that highlights the roles of cell-cell interactions during remodeling. We propose that these are fundamental to elucidating how spatiotemporal control of remodeling and coordinated circuit remodeling are achieved. We cover examples spanning various neuronal circuits in vertebrates and invertebrates and involving interactions between neurons and different cell types.
Sudarsanam S., Yaniv S., Meltzer H. & Schuldiner O.
(2020)
Journal of Cell Science.
133,
8,
jcs.232595.
The mechanisms that control intrinsic axon growth potential, and thus axon regeneration following injury, are not well understood. Developmental axon regrowth of Drosophila mushroom body γ neurons during neuronal remodeling offers a unique opportunity to study the molecular mechanisms controlling intrinsic growth potential. Motivated by the recently uncovered developmental expression atlas of γ neurons, we here focus on the role of the actin severing protein cofilin during axon regrowth. We show that Twinstar (Tsr), the fly cofilin, is a crucial regulator of both axon growth and branching during developmental remodeling of γ neurons. tsr mutant axons demonstrate growth defects both in vivo and in vitro and also exhibit actin rich filopodial-like structures at failed branch points in vivo. Our data is inconsistent with Tsr being important for increasing G-actin availability. Furthermore, analysis of microtubule localization suggests that Tsr is required for microtubule infiltration into the axon tips and branch points. Taken together, we show that Tsr promotes axon growth and branching, likely by clearing F-actin to facilitate microtubules protrusion.
Yaniv S. P., Meltzer H., Alyagor I. & Schuldiner O.
(2020)
Journal of Cell Biology.
219,
5,
201903181.
Intrinsic neurite growth potential is a key determinant of neuronal regeneration efficiency following injury. The stereotypical remodeling of Drosophila y-neurons includes developmental regrowth of pruned axons to form adult specific connections, thereby offering a unique system to uncover growth potential regulators. Motivated by the dynamic expression in remodeling y-neurons, we focus here on the role of actin elongation factors as potential regulators of developmental axon regrowth. We found that regrowth in vivo requires the actin elongation factors Ena and profilin, but not the formins that are expressed in y-neurons. In contrast, primary y-neuron sprouting in vitro requires profilin and the formin DAAM, but not Ena. Furthermore, we demonstrate that DAAM can compensate for the loss of Ena in vivo. Similarly, DAAM mutants express invariably high levels of Ena in vitro. Thus, we show that different linear actin elongation factors function in distinct contexts even within the same cell type and that they can partially compensate for each other.
Meltzer H., Marom E., Alyagor I., Mayseless O., Berkun V., Segal-Gilboa N., Unger T., Luginbuhl D. & Schuldiner O.
(2019)
Nature Communications.
10,
1,
2113.
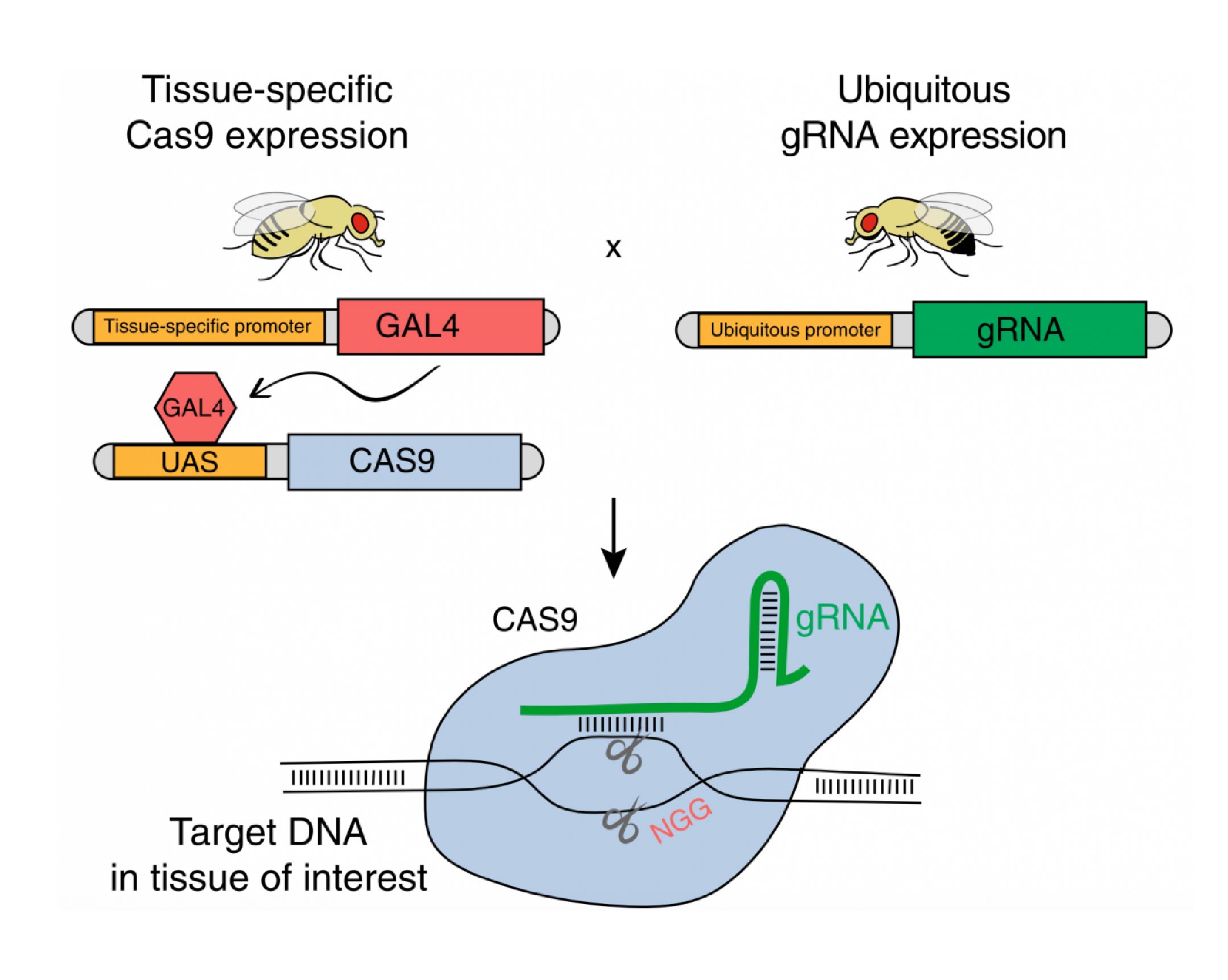
Gene editing by CRISPR/Cas9 is commonly used to generate germline mutations or perform in vitro screens, but applicability for in vivo screening has so far been limited. Recently, it was shown that in Drosophila, Cas9 expression could be limited to a desired group of cells, allowing tissue-specific mutagenesis. Here, we thoroughly characterize tissue-specific (ts)CRISPR within the complex neuronal system of the Drosophila mushroom body. We report the generation of a library of gRNA-expressing plasmids and fly lines using optimized tools, which provides a valuable resource to the fly community. We demonstrate the application of our library in a large-scale in vivo screen, which reveals insights into developmental neuronal remodeling.
Marmor-Kollet N., Gutman I., Issman-Zecharya N. & Schuldiner O.
(2019)
Frontiers in Molecular Neuroscience.
12,
232.
A fundamental question that underlies the proper wiring and function of the nervous system is how axon extension stops during development. However, our mechanistic understanding of axon stopping is currently poor. The stereotypic development of the Drosophila mushroom body (MB) provides a unique system in which three types of anatomically distinct neurons ((sic), alpha'/beta', and alpha/beta) develop and interact to form a complex neuronal structure. All three neuronal types innervate the ipsi-lateral side and do not cross the midline. Here we find that Plum, an immunoglobulin (Ig) superfamily protein that we have previously shown to function as a TGF-beta accessory receptor, is required within MB alpha/beta neurons for their midline stopping. Overexpression of Plum within MB neurons is sufficient to induce retraction of alpha/beta axons. As expected, rescue experiments revealed that Plum likely functions in alpha/beta neurons and mediates midline stopping via the downstream effector RhoGEF2. Finally, we have identified glial-derived Myoglianin (Myo) as the major TGF-beta ligand that instructs midline stopping of MB neurons. Taken together, our study strongly suggests that TGF-beta signals originating from the midline facilitate midline stopping of alpha/beta neuron in a Plum dependent manner.
Alyagor I., Berkun V., Keren-Shaul H., Marmor-Kollet N., David E., Mayseless O., Issman-Zecharya N., Amit I. & Schuldiner O.
(2018)
Developmental Cell.
47,
1,
p. 38-52
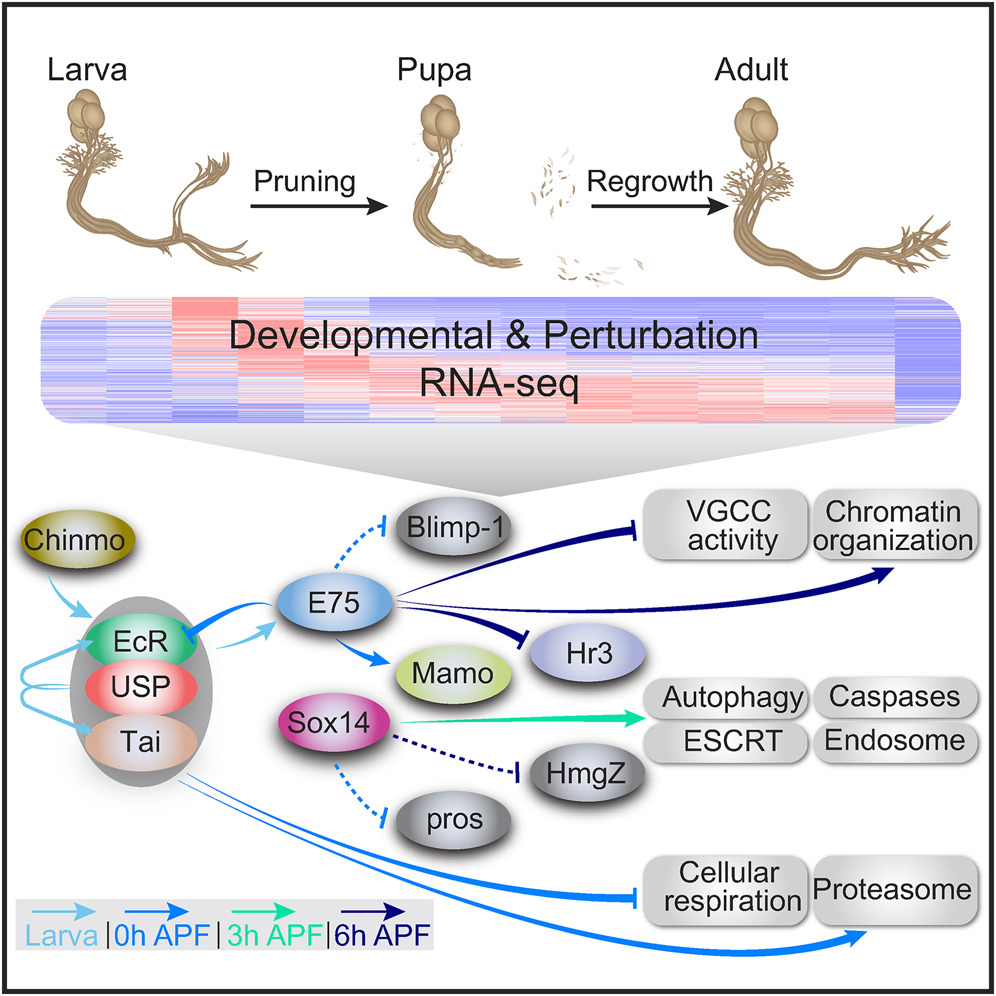
Developmental neuronal remodeling is an evolutionarily conserved mechanism required for precise wiring of nervous systems. Despite its fundamental role in neurodevelopment and proposed contribution to various neuropsychiatric disorders, the underlying mechanisms are largely unknown. Here, we uncover the fine temporal transcriptional landscape of Drosophila mushroom body γ neurons undergoing stereotypical remodeling. Our data reveal rapid and dramatic changes in the transcriptional landscape during development. Focusing on DNA binding proteins, we identify eleven that are required for remodeling. Furthermore, we sequence developing γ neurons perturbed for three key transcription factors required for pruning. We describe a hierarchical network featuring positive and negative feedback loops. Superimposing the perturbation-seq on the developmental expression atlas highlights a framework of transcriptional modules that together drive remodeling. Overall, this study provides a broad and detailed molecular insight into the complex regulatory dynamics of developmental remodeling and thus offers a pipeline to dissect developmental processes via RNA profiling. Developmental processes are often transcriptionally regulated. Alyagor et al. dissected the transcriptional landscape of a distinct neuronal type undergoing stereotypical remodeling in fine-grained temporal resolution. Superimposing this developmental expression atlas on profiles of neurons mutant for key transcription factors revealed a hierarchical regulatory network.
Mayseless O., Berns D. S., Yu X. M., Riemensperger T., Fiala A. & Schuldiner O.
(2018)
Neuron.
99,
6,
p. 1204-1215
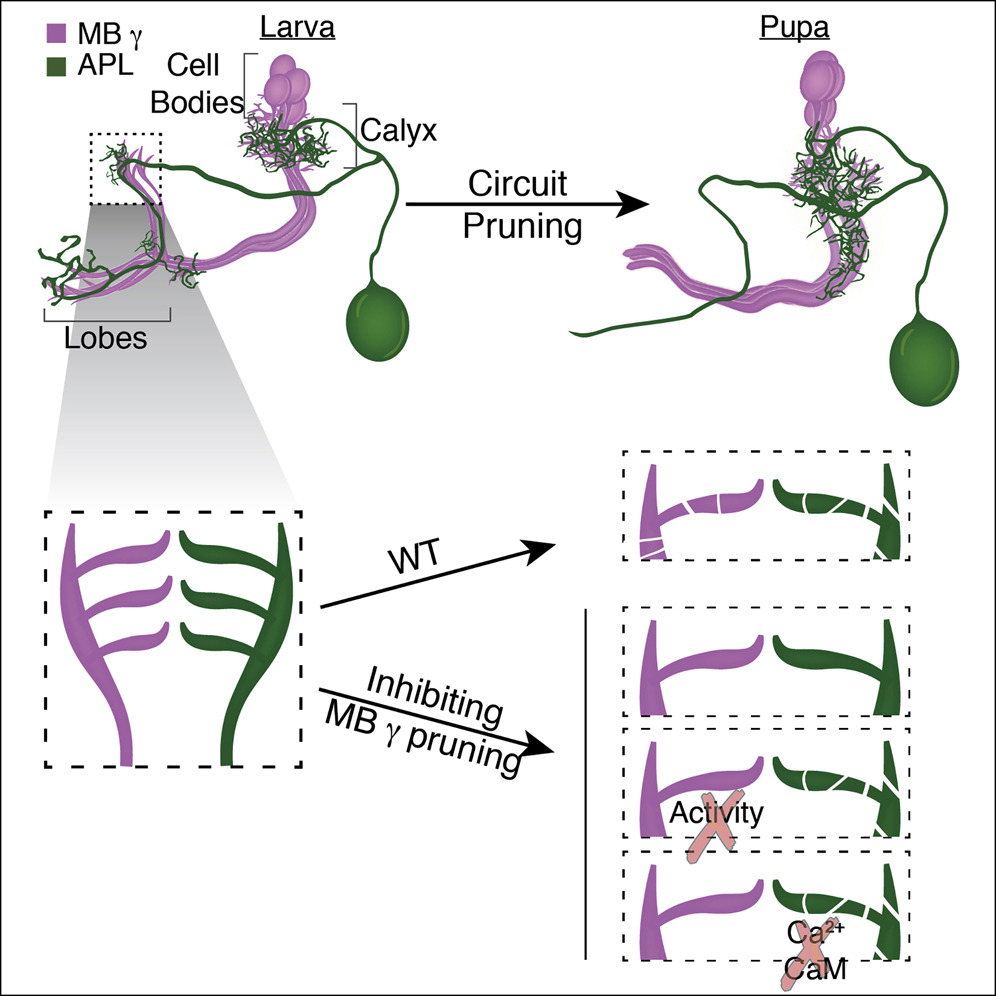
Developmental neuronal remodeling is crucial for proper wiring of the adult nervous system. While remodeling of individual neuronal populations has been studied, how neuronal circuits remodeland whether remodeling of synaptic partners is coordinatedis unknown. We found that the Drosophila anterior paired lateral (APL) neuron undergoes stereotypic remodeling during metamorphosis in a similar time frame as the mushroom body (MB) ɣ-neurons, with whom it forms a functional circuit. By simultaneously manipulating both neuronal populations, we found that cell-autonomous inhibition of ɣ-neuron pruning resulted in the inhibition of APL pruning in a process that is mediated, at least in part, by Ca
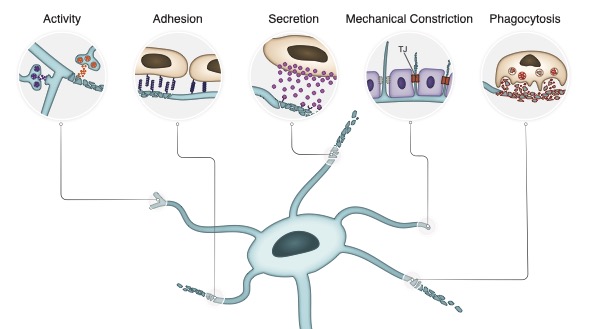
2+-Calmodulin and neuronal activity dependent interaction. Finally, ectopic unpruned MB ɣ axons display ectopic connections with the APL, as well as with other neurons, at the adult, suggesting that inhibiting remodeling of one neuronal type can affect the functional wiring of the entire micro-circuit. Developmental remodeling is crucial for stereotypic neural circuit formation. Mayseless et al. describe a neural circuit in Drosophila that undergoes coordinated remodeling. They show that neurons within the same circuit interact via neuronal activity and Ca
2+/Calmodulin signaling to coordinate remodeling.
Meltzer H. & Schuldiner O.
(2016)
Neuron.
92,
4,
p. 677-680
In this issue of Neuron, Brill et al. (2016) demonstrate that, during synapse elimination in the developing neuromuscular junction, branch-specific microtubule destabilization results in arrested axonal transport and induces axon branch loss. This process is mediated in part by the neurodegeneration-associated, microtubule-severing protein spastin.
Yaniv S. P. & Schuldiner O.
(2016)
Wiley interdisciplinary reviews. Developmental biology.
5,
5,
p. 618-635
Developmental neuronal remodeling is a crucial step in sculpting the final and mature brain connectivity in both vertebrates and invertebrates. Remodeling includes degenerative events, such as neurite pruning, that may be followed by regeneration to form novel connections during normal development. Drosophila provides an excellent model to study both steps of remodeling since its nervous system undergoes massive and stereotypic remodeling during metamorphosis. Although pruning has been widely studied, our knowledge of the molecular and cellular mechanisms is far from complete. Our understanding of the processes underlying regrowth is even more fragmentary. In this review, we discuss recent progress by focusing on three groups of neurons that undergo stereotypic pruning and regrowth during metamorphosis, the mushroom body γ neurons, the dendritic arborization neurons and the crustacean cardioactive peptide peptidergic neurons. By comparing and contrasting the mechanisms involved in remodeling of these three neuronal types, we highlight the common themes and differences as well as raise key questions for future investigation in the field. WIREs Dev Biol 2016, 5:618635. doi: 10.1002/wdev.241. For further resources related to this article, please visit the WIREs website.
Yaron A. & Schuldiner O.
(2016)
Current Biology.
26,
13,
p. R628-R639
Cell death is an inherent process that is required for the proper wiring of the nervous system. Studies over the last four decades have shown that, in a parallel developmental pathway, axons and dendrites are eliminated without the death of the neuron. This developmentally regulated `axonal death' results in neuronal remodeling, which is an essential mechanism to sculpt neuronal networks in both vertebrates and invertebrates. Studies across various organisms have demonstrated that a conserved strategy in the formation of adult neuronal circuitry often involves generating too many connections, most of which are later eliminated with high temporal and spatial resolution. Can neuronal remodeling be regarded as developmentally and spatially regulated neurodegeneration? It has been previously speculated that injury-induced degeneration ( Wallerian degeneration) shares some molecular features with `dying back' neurodegenerative diseases. In this opinion piece, we examine the similarities and differences between the mechanisms regulating neuronal remodeling and those being perturbed in dying back neurodegenerative diseases. We focus primarily on amyotrophic lateral sclerosis and peripheral neuropathies and highlight possible shared pathways and mechanisms. While mechanistic data are only just beginning to emerge, and despite the inherent differences between disease-oriented and developmental processes, we believe that some of the similarities between these developmental and disease-initiated degeneration processes warrant closer collaborations and crosstalk between these different fields.
Marmor-Kollet N. & Schuldiner O.
(2016)
Developmental Neurobiology.
76,
3,
p. 262-276
The molecular mechanisms regulating intrinsic axon growth potential during development or following injury remain largely unknown despite their vast importance. Here, we have established a neurite sprouting assay of primary cultured mushroom body (MB) neurons. We used the MARCM technique to both mark and manipulate MB neurons, enabling us to quantify the sprouting abilities of single WT and mutant neurons originating from flies at different developmental stages. Sprouting of dissociated MB neurons was dependent on wnd, the DLK ortholog, a conserved gene that is required for axon regeneration. Next, and as expected, we found that the sprouting ability of adult MB neurons was significantly decreased. In contrast, and to our surprise, we found that pupal-derived neurons exhibit increased sprouting compared with neurons derived from larvae, suggesting the existence of an elevated growth potential state. We then contrasted the molecular requirements of neurite sprouting to developmental axon regrowth of MB γ neurons, a process that we have previously shown requires the nuclear receptor UNF acting via the target of rapamycin (TOR) pathway. Strikingly, we found that while TOR was required for neurite sprouting, UNF was not. In contrast, we found that PTEN inhibits sprouting in adult neurons, suggesting that TOR is regulated by the PI3K/PTEN pathway during sprouting and by UNF during developmental regrowth. Interestingly, the PI3K pathway as well as Wnd were not required for developmental regrowth nor for initial axon outgrowth suggesting that axon growth during circuit formation, remodeling, and regeneration share some molecular components but differ in others.
Rabinovich D., Yaniv S. P., Alyagor I. & Schuldiner O.
(2016)
Cell.
164,
1-2,
p. 170-182
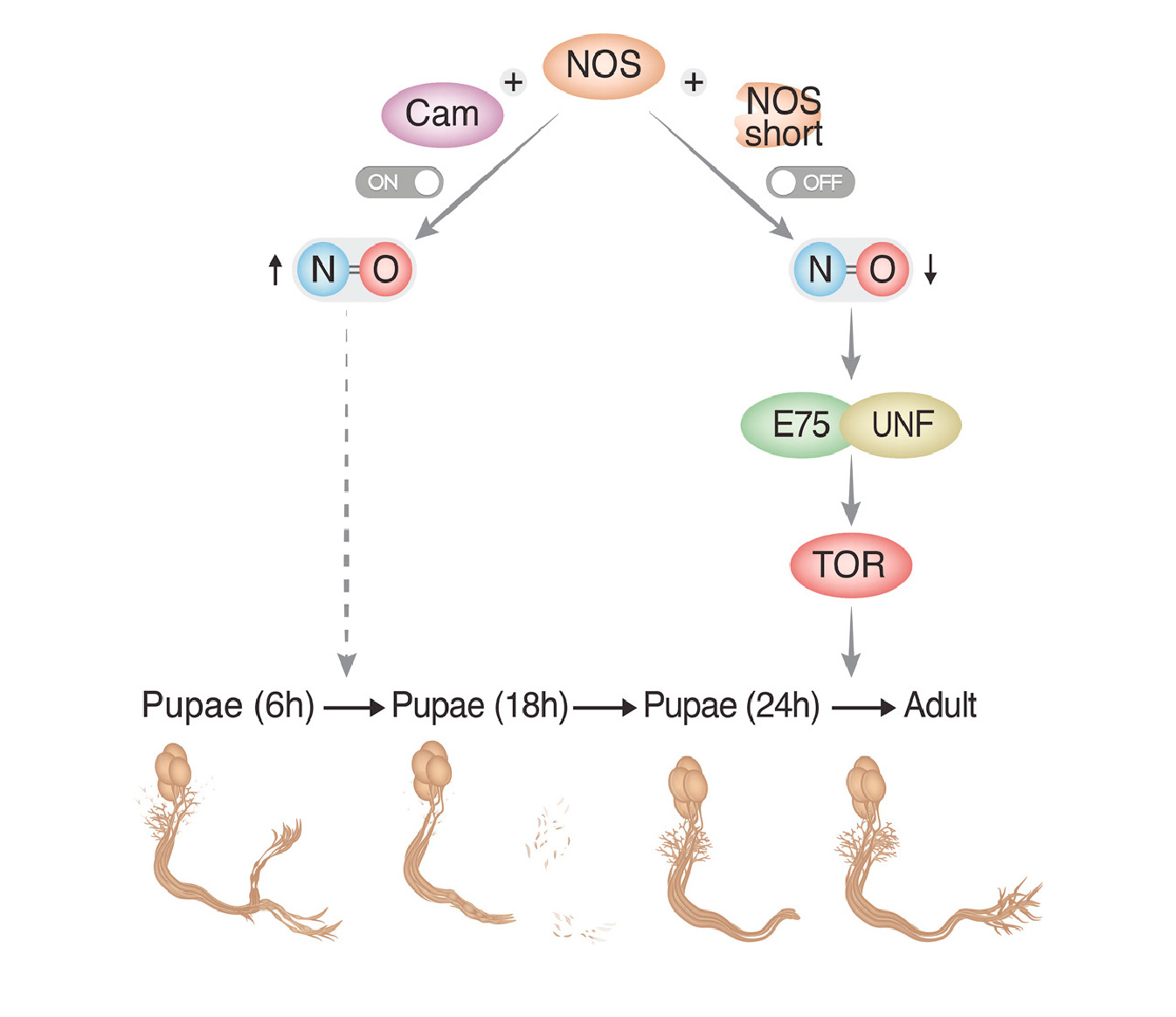
During development, neurons switch among growth states, such as initial axon outgrowth, axon pruning, and regrowth. By studying the stereotypic remodeling of the Drosophila mushroom body (MB), we found that the heme-binding nuclear receptor E75 is dispensable for initial axon outgrowth of MB γ neurons but is required for their developmental regrowth. Genetic experiments and pharmacological manipulations on ex-vivo-cultured brains indicate that neuronally generated nitric oxide (NO) promotes pruning but inhibits regrowth. We found that high NO levels inhibit the physical interaction between the E75 and UNF nuclear receptors, likely accounting for its repression of regrowth. Additionally, NO synthase (NOS) activity is downregulated at the onset of regrowth, at least partially, by short inhibitory NOS isoforms encoded within the NOS locus, indicating how NO production could be developmentally regulated. Taken together, these results suggest that NO signaling provides a switching mechanism between the degenerative and regenerative states of neuronal remodeling.
Bornstein B., Zahavi E. E., Gelley S., Zoosman M., Yaniv S. P., Fuchs O., Porat Z., Perlson E. & Schuldiner O.
(2015)
Neuron.
88,
5,
p. 926-940
Developmental axon pruning is essential for normal brain wiring in vertebrates and invertebrates. How axon pruning occurs in vivo is not well understood. In a mosaic loss-of-function screen, we found that Bsk, the Drosophila JNK, is required for axon pruning of mushroom body γ neurons, but not their dendrites. By combining in vivo genetics, biochemistry, and high-resolution microscopy, we demonstrate that the mechanism by which Bsk is required for pruning is through reducing the membrane levels of the adhesion molecule Fasciclin II (FasII), the NCAM ortholog. Conversely, overexpression of FasII is sufficient to inhibit axon pruning. Finally, we show that overexpressing other cell adhesion molecules, together with weak attenuation of JNK signaling, strongly inhibits pruning. Taken together, we have uncovered a novel and unexpected interaction between the JNK pathway and cell adhesion and found that destabilization of cell adhesion is necessary for efficient pruning.
Rabinovich D., Mayseless O. & Schuldiner O.
(2015)
FRONTIERS IN CELLULAR NEUROSCIENCE.
9,
AUGUST,
327.
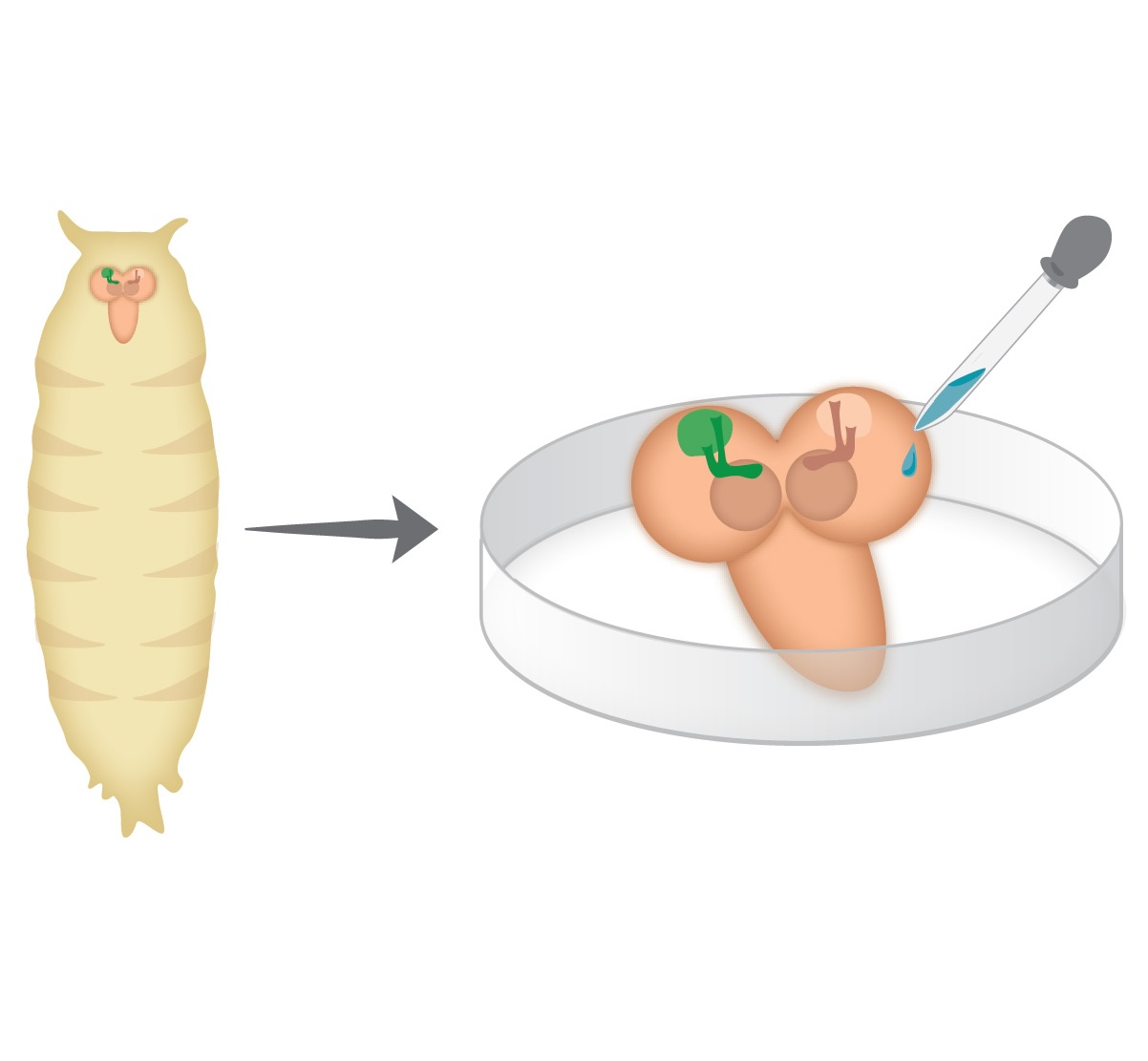
Holometabolous insects, including Drosophila melanogaster, undergo complete metamorphosis that includes a pupal stage. During metamorphosis, the Drosophila nervous system undergoes massive remodeling and growth, that include cell death and large-scale axon and synapse elimination as well as neurogenesis, developmental axon regrowth, and formation of new connections. Neuronal remodeling is an essential step in the development of vertebrate and invertebrate nervous systems. Research on the stereotypic remodeling of Drosophila mushroom body (MB) γ neurons has contributed to our knowledge of the molecular mechanisms of remodeling but our knowledge of the cellular mechanisms remain poorly understood. A major hurdle in understanding various dynamic processes that occur during metamorphosis is the lack of time-lapse resolution. The pupal case and opaque fat bodies that enwrap the central nervous system (CNS) make live-imaging of the central brain in-vivo impossible. We have established an ex vivo long-term brain culture system that supports the development and neuronal remodeling of pupal brains. By optimizing culture conditions and dissection protocols, we have observed development in culture at kinetics similar to what occurs in vivo. Using this new method, we have obtained the first time-lapse sequence of MB γ neurons undergoing remodeling in up to a single cell resolution. We found that axon pruning is initiated by blebbing, followed by one-two nicks that seem to initiate a more widely spread axon fragmentation. As such, we have set up some of the tools and methodologies needed for further exploration of the cellular mechanisms of neuronal remodeling, not limited to the MB. The long-term ex vivo brain culture system that we report here could be used to study dynamic aspects of neurodevelopment of any Drosophila neuron.
Schuldiner O. & Yaron A.
(2015)
Cellular and Molecular Life Sciences.
72,
1,
p. 101-119
The precise wiring of the nervous system is a combined outcome of progressive and regressive events during development. Axon guidance and synapse formation intertwined with cell death and neurite pruning sculpt the mature circuitry. It is now well recognized that pruning of dendrites and axons as means to refine neuronal networks, is a wide spread phenomena required for the normal development of vertebrate and invertebrate nervous systems. Here we will review the arising principles of cellular and molecular mechanisms of neurite pruning. We will discuss these principles in light of studies in multiple neuronal systems, and speculate on potential explanations for the emergence of neurite pruning as a mechanism to sculpt the nervous system.
Issman-Zecharya N. & Schuldiner O.
(2014)
Developmental Cell.
31,
4,
p. 461-473
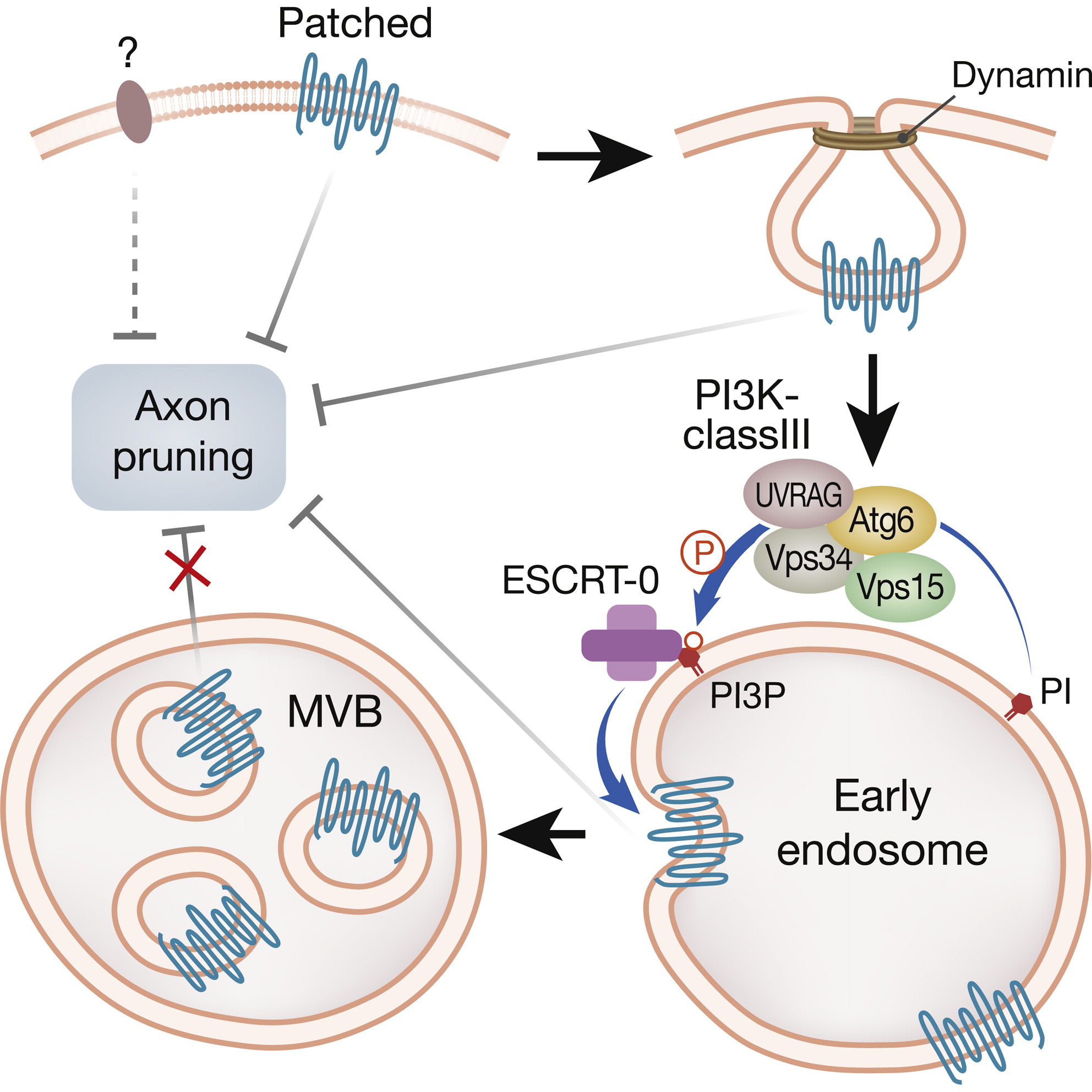
Developmental axon pruning is essential for wiring the mature nervous system, but its regulation remains poorly understood. Here we show that the endosomal-lysosomal pathway regulates developmental pruning of Drosophila mushroom body γ neurons. We demonstrate that the UV radiation resistance-associated gene (UVRAG) functions together with all core components of the phosphatidylinositol 3-kinase class III (PI3K-cIII) complex to promote pruning via the endocytic pathway. By studying several PI(3)P binding proteins, we found that Hrs, a subunit of the ESCRT-0 complex, required for multivesicular body (MVB) maturation, is essential for normal pruning progression. Thus, we hypothesized the existence of an inhibitory signal that needs to be downregulated. Finally, our data suggest that the Hedgehog receptor, Patched, is the source of this inhibitory signal likely functioning in a Smo-independent manner. Taken together, our invivo study demonstrates that the PI3K-cIII complex is essential for downregulating Patched via the endosomal-lysosomal pathway to execute axon pruning.
Yu F. & Schuldiner O.
(2014)
Current Opinion in Neurobiology.
27,
p. 192-198
Pruning, a process by which neurons selectively remove exuberant or unnecessary processes without causing cell death, is crucial for the establishment of mature neural circuits during animal development. Yet relatively little is known about molecular and cellular mechanisms that govern neuronal pruning. Holometabolous insects, such as Drosophila, undergo complete metamorphosis and their larval nervous systems are replaced with adult-specific ones, thus providing attractive models for studying neuronal pruning. Drosophila mushroom body and dendritic arborization neurons have been utilized as two appealing systems to elucidate the underlying mechanisms of axon and dendrite pruning, respectively. In this review we highlight recent developments and discuss some similarities and differences in the mechanisms that regulate these two distinct modes of neuronal pruning in Drosophila.
Hakim Y., Yaniv S. P. & Schuldiner O.
(2014)
PLoS ONE.
9,
1,
e86178.
Axon pruning is an evolutionarily conserved strategy used to remodel neuronal connections during development. The Drosophila mushroom body (MB) undergoes neuronal remodeling in a highly stereotypical and tightly regulated manner, however many open questions remain. Although it has been previously shown that glia instruct pruning by secreting a TGF-β ligand, myoglianin, which primes MB neurons for fragmentation and also later engulf the axonal debris once fragmentation has been completed, which glia subtypes participate in these processes as well as the molecular details are unknown. Here we show that, unexpectedly, astrocytes are the major glial subtype that is responsible for the clearance of MB axon debris following fragmentation, even though they represent only a minority of glia in the MB area during remodeling. Furthermore, we show that astrocytes both promote fragmentation of MB axons as well as clear axonal debris and that this process is mediated by ecdysone signaling in the astrocytes themselves. In addition, we found that blocking the expression of the cell engulfment receptor Draper in astrocytes only affects axonal debris clearance. Thereby we uncoupled the function of astrocytes in promoting axon fragmentation to that of clearing axonal debris after fragmentation has been completed. Our study finds a novel role for astrocytes in the MB and suggests two separate pathways in which they affect developmental axon pruning.
Yu X. M., Gutman I., Mosca T. J., Iram T., Oezkan E., Garcia K. C., Luo L. & Schuldiner O.
(2013)
Neuron.
78,
3,
p. 456-468
Axon pruning during development is essential for proper wiring of the mature nervous system, but itsregulation remains poorly understood. We have identified an immunoglobulin superfamily (IgSF) transmembrane protein, Plum, that is cell autonomously required for axon pruning of mushroom body (MB) γ neurons and for ectopic synapse refinement at the developing neuromuscular junction in. Drosophila. Plum promotes MB γ neuron axon pruning by regulating the expression of Ecdysone Receptor-B1, a key initiator of axon pruning. Genetic analyses indicate that Plum acts to facilitate signaling of Myoglianin, a glial-derived TGF-β, on MB γ neurons upstream of the type-I TGF-β receptor Baboon. Myoglianin, Baboon, and Ecdysone Receptor-B1 are also required for neuromuscular junction ectopic synapse refinement. Our study highlights both IgSF proteins and TGF-β facilitation as key promoters of developmental axon elimination and demonstrates a mechanistic conservation between MB axon pruning during metamorphosis and the refinement of ectopic larval neuromuscular connections
Yaniv S. P., Issman-Zecharya N., Oren-Suissa M., Podbilewicz B. & Schuldiner O.
(2012)
Current Biology.
22,
19,
p. 1774-1782
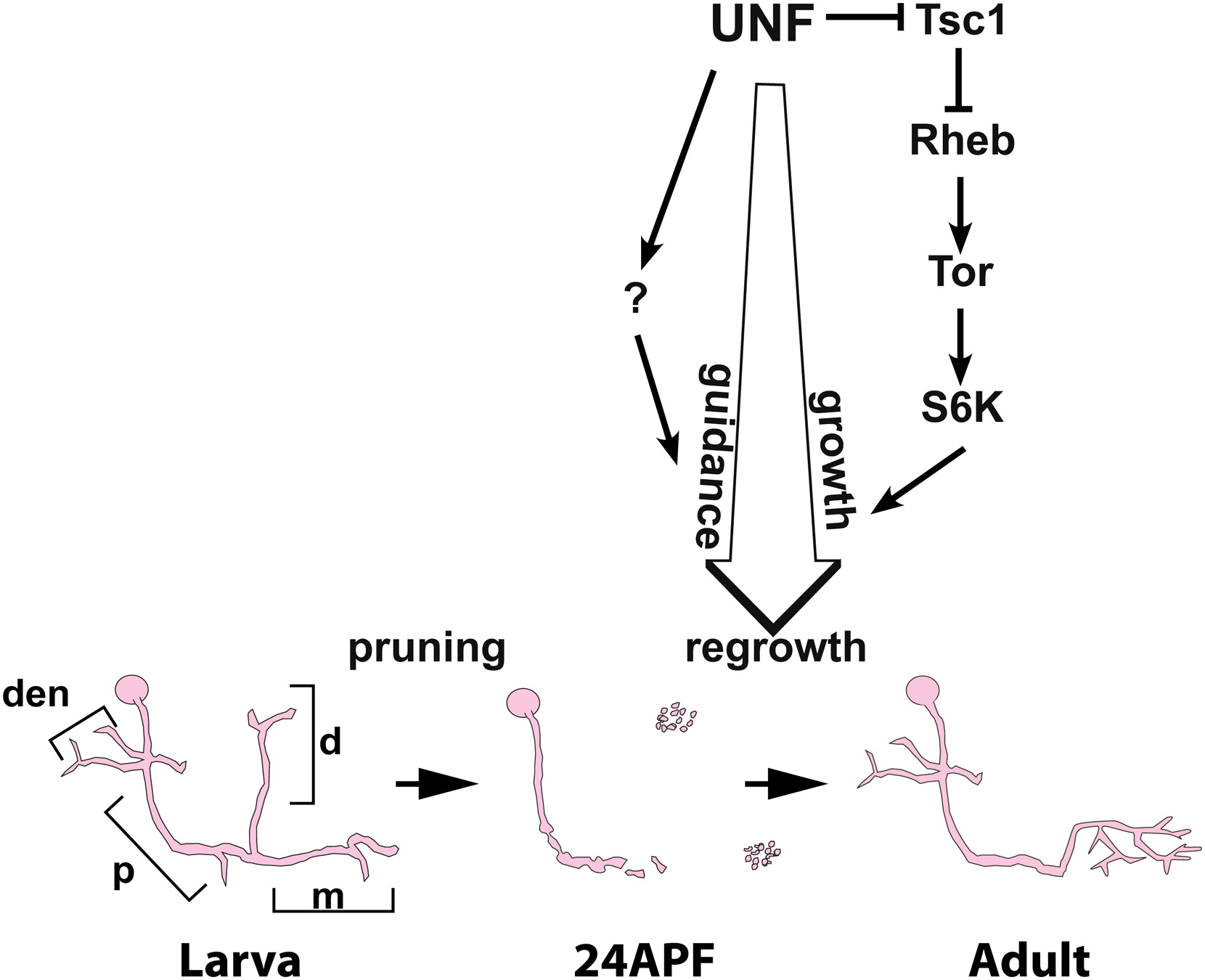
Background: The molecular mechanisms that determine axonal growth potential are poorly understood. Intrinsic growth potential decreases with age, and thus one strategy to identify molecular pathways controlling intrinsic growth potential is by studying developing young neurons. The programmed and stereotypic remodeling of Drosophila mushroom body (MB) neurons during metamorphosis offers a unique opportunity to uncover such mechanisms. Despite emerging insights into MB γ-neuron axon pruning, nothing is known about the ensuing axon re-extension. Results: Using mosaic loss of function, we found that the nuclear receptor UNF (Nr2e3) is cell autonomously required for the re-extension of MB γ-axons following pruning, but not for the initial growth or guidance of any MB neuron type. We found that UNF promotes this process of developmental axon regrowth via the TOR pathway as well as a late axon guidance program via an unknown mechanism. We have thus uncovered a novel developmental program of axon regrowth that is cell autonomously regulated by the UNF nuclear receptor and the TOR pathway. Conclusions: Our results suggest that UNF activates neuronal re-extension during development. Taken together, we show that axon growth during developmental remodeling is mechanistically distinct from initial axon outgrowth. Due to the involvement of the TOR pathway in axon regeneration following injury, our results also suggests that developmental regrowth shares common molecular mechanisms with regeneration following injury.
Schuldiner O., Berdnik D., Levy J. M., Wu J. S., Luginbuhl D., Gontang A. C. & Luo L.
(2008)
Developmental Cell.
14,
2,
p. 227-238
Developmental axon pruning is widely used to refine neural circuits. We performed a mosaic screen to identify mutations affecting axon pruning of Drosophila mushroom body γ neurons. We constructed a modified piggyBac vector with improved mutagenicity and generated insertions in >2000 genes. We identified two cohesin subunits (SMC1 and SA) as being essential for axon pruning. The cohesin complex maintains sister-chromatid cohesion during cell division in eukaryotes. However, we show that the pruning phenotype in SMC1-/- clones is rescued by expressing SMC1 in neurons, revealing a postmitotic function. SMC1-/- clones exhibit reduced levels of the ecdysone receptor EcR-B1, a key regulator of axon pruning. The pruning phenotype is significantly suppressed by overexpressing EcR-B1 and is enhanced by a reduced dose of EcR, supporting a causal relationship. We also demonstrate a postmitotic role for SMC1 in dendrite targeting of olfactory projection neurons. We suggest that cohesin regulates diverse aspects of neuronal morphogenesis.