
Our group was the first to show a relationship between allostery in GroEL and GroEL-assisted protein folding rates (Yifrach and Horovitz, 2000). Using different cooperativity mutants of GroEL, we found a linear relationship between the folding rate of mouse dihydrofolate reductase (under conditions where its folding is GroEL-dependent but GroES-independent) and the rate and Hill coefficient of the T→R allosteric transition.
We also analyzed the importance of GroEL allostery in vivo by generating E. coli strains that express only plasmid-derived GroEL (wild-type or different mutants which are defective in their allosteric properties) and characterizing their phenotypes (Fridmann et al., 2000).
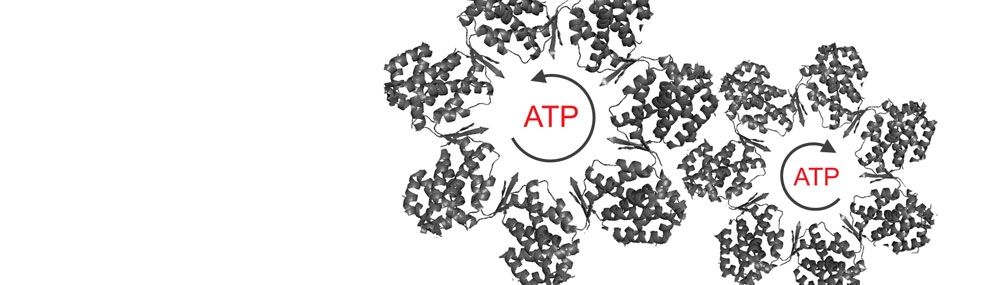
More recently, we have been interested in investigating the impact of the mechanism of allosteric switching (concerted vs. sequential) on the folding function of chaperonins. Our working hypothesis is that sequential conformational changes facilitate domain-by-domain folding and, thus, enable CCT/TRiC to better assist folding of multi-domain proteins that are more common in eukaryotes. By contrast, the concerted allosteric transitions of GroEL facilitate protein substrate release in an all-or-none manner folding, thereby enhancing the folding efficiency of single-domain proteins that are more common in prokaryotes. In the case of GroEL, this hypothesis has been tested experimentally and confirmed (Kipnis et al., 2007; Papo et al., 2008). Work is underway to test this hypothesis for CCT/TRiC .