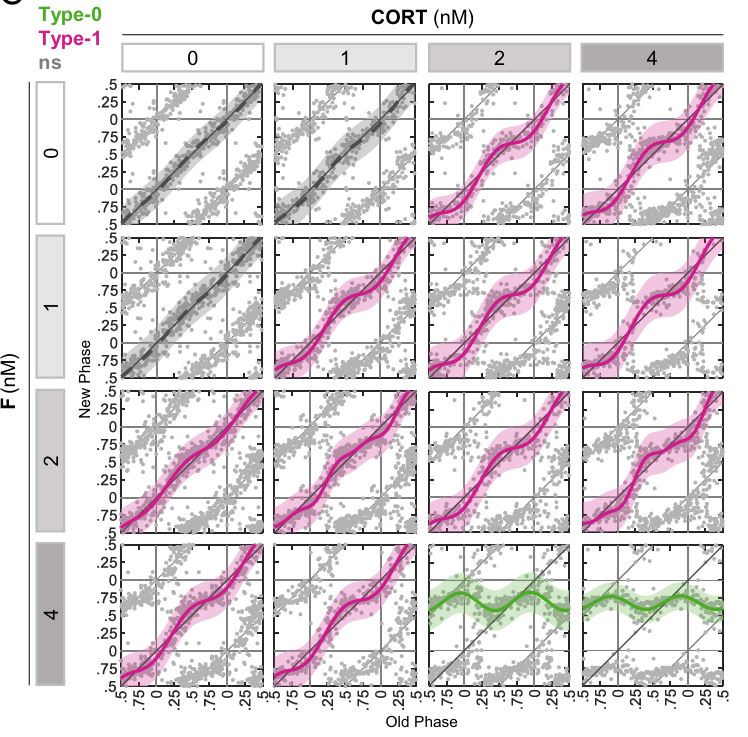
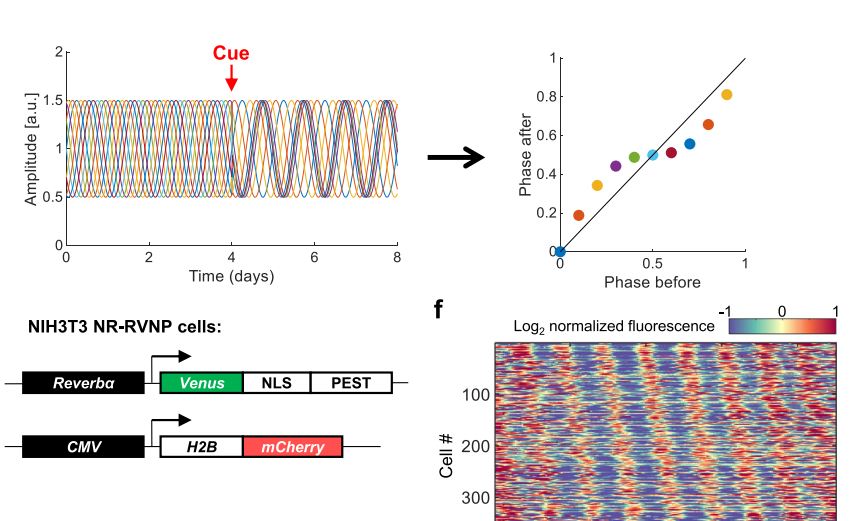
Circadian clocks are cell-autonomous oscillators that are present in most cells of the body and temporally coordinate their function. Alignment of cellular clocks with each other and the environment is mediated mostly through blood borne signals. Although serum is a potent resetting signal for circadian clocks, the underlying intracellular molecular underpinnings are largely unknown. Here, we employ Circa-SCOPE, a high-throughput single-cell method for constructing Phase Transition Curves (PTCs), to classify intracellular signaling pathways and clock-components that participate in clock resetting by serum. We identify steroid hormone, including sex-hormone receptors as key mediators of serum-induced phase resetting. Unexpectedly, we discover that Cry2 plays a central role in the response to serum and specifically to steroid hormones, irrespectively of its effect on the clock period-length. Furthermore, we find that PTCs are largely unaffected by the period-length. Overall, our findings provide important insight on intracellular determinant of the clock response to serum.
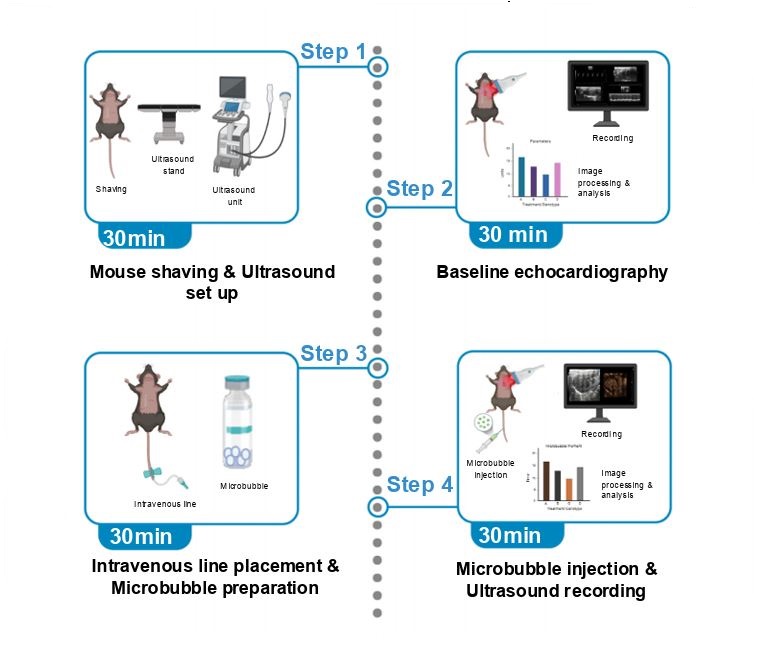
Monitoring intrapulmonary vasodilation in animal models is vital for understanding pulmonary vascular physiology and disease mechanisms. Here, we present a protocol for the real-time visualization and quantification of pulmonary microvascular dynamics in living mice using contrast echocardiography alongside microbubble injection. We describe steps for mouse preparation, baseline echocardiography, microbubble administration, and imaging to capture dynamic changes in pulmonary vasculature. Closely resembling clinical technique used in humans, this methodology provides an effective tool for investigating the pathophysiology of diverse cardio-pulmonary vascular diseases. For complete details on the use and execution of this protocol, please refer to Dandavate et al.1
Deota S., Pendergast J. S., Kolthur-Seetharam U., Esser K. A., Gachon F., Asher G., Dibner C., Benitah S. A., Escobar C., Muoio D. M., Zhang E. E., Hotamışlıgil G. S., Bass J., Takahashi J. S., Rabinowitz J. D., Lamia K. A., de Cabo R., Kajimura S., Longo V. D., Xu Y., Lazar M. A., Verdin E., Zierath J. R., Auwerx J., Drucker D. J. & Panda S.
(2025)
Nature metabolism.
7,
3,
p. 454-468
142.
The constant expansion of the field of metabolic research has led to more nuanced and sophisticated understanding of the complex mechanisms that underlie metabolic functions and diseases. Collaborations with scientists of various fields such as neuroscience, immunology and drug discovery have further enhanced the ability to probe the role of metabolism in physiological processes. However, many behaviours, endocrine and biochemical processes, and the expression of genes, proteins and metabolites have daily ~24-h biological rhythms and thus peak only at specific times of the day. This daily variation can lead to incorrect interpretations, lack of reproducibility across laboratories and challenges in translating preclinical studies to humans. In this Review, we discuss the biological, environmental and experimental factors affecting circadian rhythms in rodents, which can in turn alter their metabolic pathways and the outcomes of experiments. We recommend that these variables be duly considered and suggest best practices for designing, analysing and reporting metabolic experiments in a circadian context.
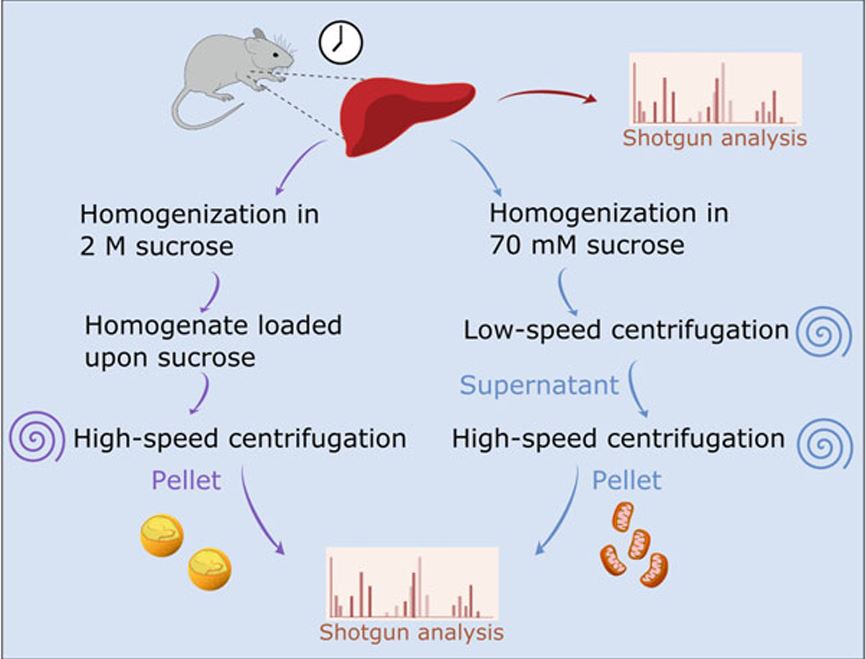
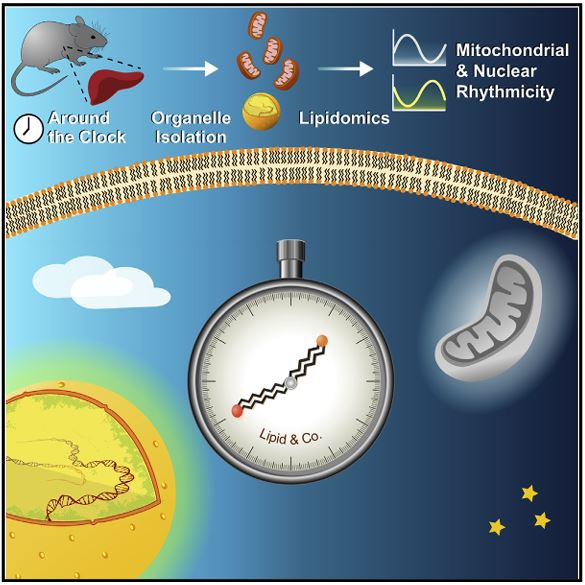
Lipidomic methodologies, in conjunction with biochemical isolation of intracellular organelles such as the nucleus, have identified a wide variety of nuclear lipid species. The nuclear lipid composition, for example, exhibits daily changes and is controlled both by the circadian clock and feeding behavior. However, the functions of many of these lipids in the nucleus remain elusive. In this chapter we describe in detail the procedure for nuclear lipidomics; from biochemical isolation of nuclei from mouse liver and cultured cells to the unbiased lipid identification and quantification using Multi-Dimensional Mass Spectrometry Shotgun Lipidomics (MDMS-SL). The approach described herein is applicable with some modifications for different tissues and various animal models, as well as a wide variety of cultured cells. It is expected to yield new insights into various aspects of nuclear lipid composition and function in the future.
Dandavate V., Bolshette N., Van Drunen R., Manella G., Bueno-Levy H., Zerbib M., Kawano I., Golik M., Adamovich Y. & Asher G.
(2024)
Cell Metabolism.
36,
9,
p. 2038-2053.e5
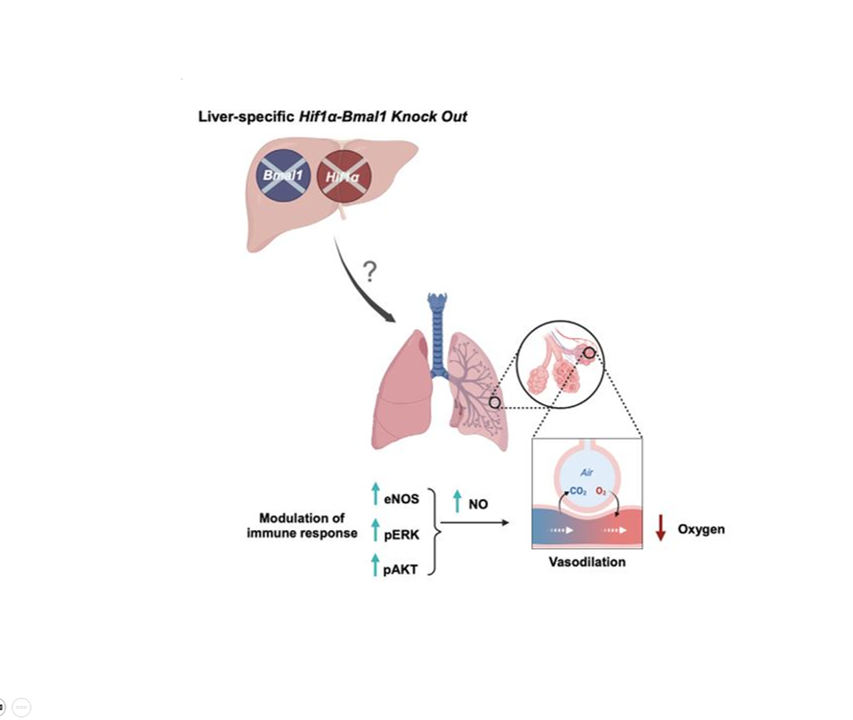
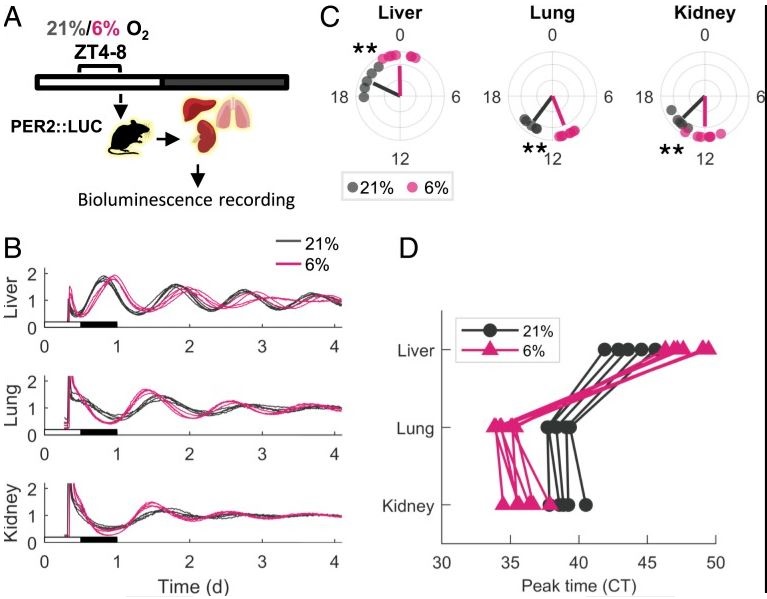
The transcriptional response to hypoxia is temporally regulated, yet the molecular underpinnings and physiological implications are unknown. We examined the roles of hepatic Bmal1 and Hif1α in the circadian response to hypoxia in mice. We found that the majority of the transcriptional response to hypoxia is dependent on either Bmal1 or Hif1α, through shared and distinct roles that are daytime determined. We further show that hypoxia-inducible factor (HIF)1α accumulation upon hypoxia is temporally regulated and Bmal1 dependent. Unexpectedly, mice lacking both hepatic Bmal1 and Hif1α are hypoxemic and exhibit increased mortality upon hypoxic exposure in a daytime-dependent manner. These mice display mild liver dysfunction with pulmonary vasodilation likely due to extracellular signaling regulated kinase (ERK) activation, endothelial nitric oxide synthase, and nitric oxide accumulation in lungs, suggestive of hepatopulmonary syndrome. Our findings indicate that hepatic BMAL1 and HIF1α are key time-dependent regulators of the hypoxic response and can provide molecular insights into the pathophysiology of hepatopulmonary syndrome.
Bolshette N., Ezagouri S., Dandavate V., Karavaeva I., Golik M., Wang H., Espenshade P. J., Osborne T. F., Han X. & Asher G.
(2023)
PLoS Biology.
21,
11,
e3002367.
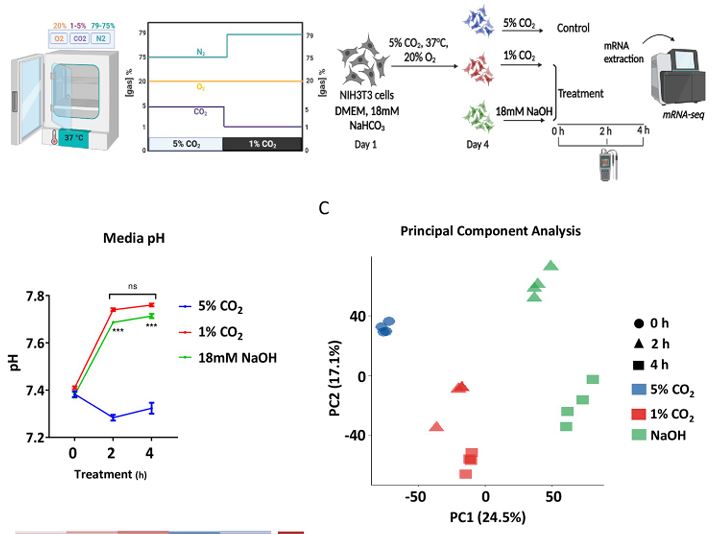
AU In mammals,: Pleaseconfirmthatallheadinglevelsarerepresentedcorrectly O2 and CO2 levels are tightly regulated and are:altered under various pathological conditions. While the molecular mechanisms that participate in O2 sensing are well characterized, little is known regarding the signaling pathways that participate in CO2 signaling and adaptation. Here, we show that CO2 levels control a distinct cellular transcriptional response that differs from mere pH changes. Unexpectedly, we discovered that CO2 regulates the expression of cholesterogenic genes in a SREBP2-dependent manner and modulates cellular cholesterol accumulation. Molecular dissection of the underlying mechanism suggests that CO2 triggers SREBP2 activation through changes in endoplasmic reticulum (ER) membrane cholesterol levels. Collectively, we propose that SREBP2 participates in CO2 signaling and that cellular cholesterol levels can be modulated by CO2 through SREBP2.
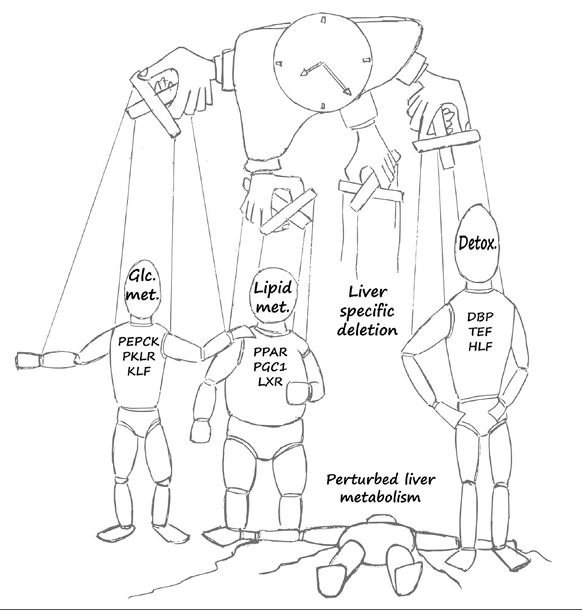
A wide variety of liver functions are regulated daily by the liver circadian clock and via systemic circadian control by other organs and cells within the gastrointestinal tract as well as the microbiome and immune cells. Disruption of the circadian system, as occurs during jetlag, shift work or an unhealthy lifestyle, is implicated in several liver-related pathologies, ranging from metabolic diseases such as obesity, type 2 diabetes mellitus and nonalcoholic fatty liver disease to liver malignancies such as hepatocellular carcinoma. In this Review, we cover the molecular, cellular and organismal aspects of various liver pathologies from a circadian viewpoint, and in particular how circadian dysregulation has a role in the development and progression of these diseases. Finally, we discuss therapeutic and lifestyle interventions that carry health benefits through support of a functional circadian clock that acts in synchrony with the environment.
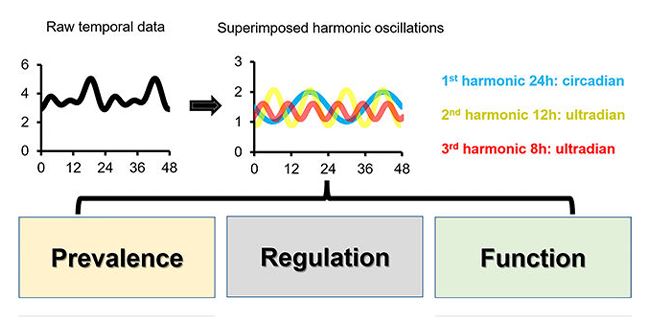
The mammalian liver must cope with various metabolic and physiological changes that normally recur every day and primarily stem from daily cycles of rest-activity and fasting-feeding. Although a large body of evidence supports the reciprocal regulation of circadian rhythms and liver function, the research on the hepatic ultradian rhythms have largely been lagging behind. However, with the advent of more cost-effective high-throughput omics technologies, high-resolution time-lapse imaging, and more robust and powerful mathematical tools, several recent studies have shed new light on the presence and functions of hepatic ultradian rhythms. In this review, we will first very briefly discuss the basic principles of circadian rhythms, and then cover in greater details the recent literature related to ultradian rhythms. Specifically, we will highlight the prevalence and mechanisms of hepatic 12-h rhythms, and 8-h rhythms, which cycle at the second and third harmonics of circadian frequency. Finally, we also refer to ultradian rhythms with other frequencies and examine the limitations of the current approaches as well as the challenges related to identifying ultradian rhythm and addressing their molecular underpinnings.
Manella G., Bolshette N., Golik M. & Asher G.
(2022)
Proceedings of the National Academy of Sciences of the United States of America.
119,
44,
e220993311.
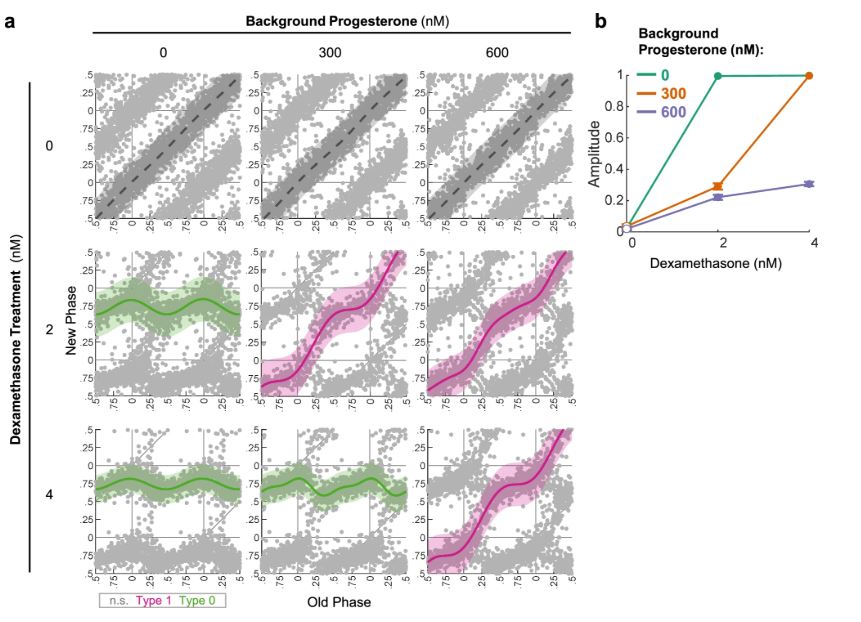
Circadian clocks are synchronized by external timing cues to align with one another and the environment. Various signaling pathways have been shown to independently reset the phase of the clock. However, in the body, circadian clocks are exposed to a multitude of potential timing cues with complex temporal dynamics, raising the question of how clocks integrate information in response to multiple signals. To investigate different modes of signal integration by the circadian clock, we used Circa-SCOPE, a method we recently developed for high-throughput phase resetting analysis. We found that simultaneous exposure to different combinations of known pharmacological resetting agents elicits a diverse range of responses. Often, the response was nonadditive and could not be readily predicted by the response to the individual signals. For instance, we observed that dexamethasone is dominant over other tested inputs. In the case of signals administered sequentially, the background levels of a signal attenuated subsequent resetting by the same signal, but not by signals acting through a different pathway. This led us to examine whether the circadian clock is sensitive to relative rather than absolute levels of the signal. Importantly, our analysis revealed the involvement of a signal-specific fold-change detection mechanism in the clock response. This mechanism likely stems from properties of the signaling pathway that are upstream to the clock. Overall, our findings elucidate modes of input integration by the circadian clock, with potential relevance to clock resetting under both physiological and pathological conditions.
Manella G., Ezagouri S., Champigneulle B., Gaucher J., Mendelson M., Lemarie E., Stauffer E., Pichon A., Howe C. A., Doutreleau S., Golik M., Verges S. & Asher G.
(2022)
Cell Reports.
40,
7,
111213.
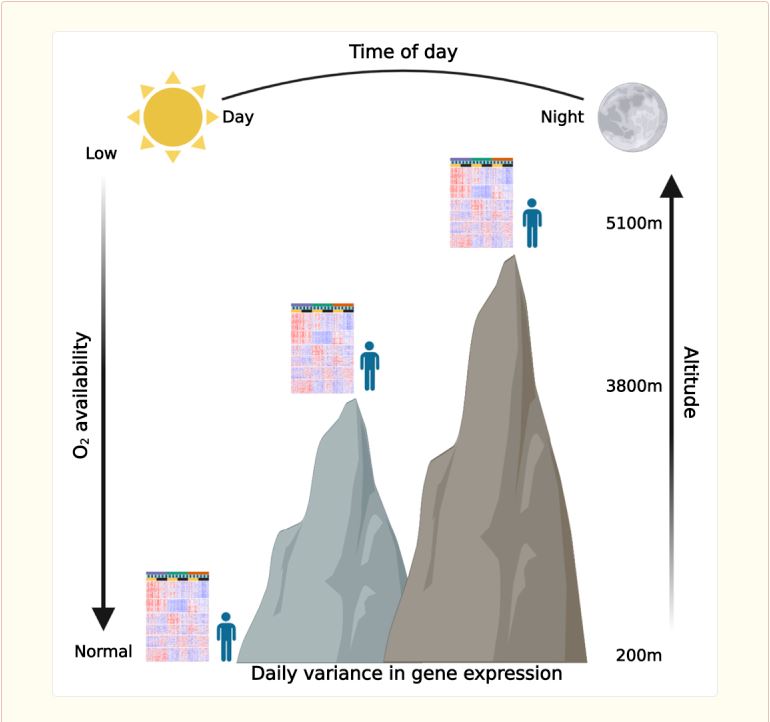
High altitude exposes humans to hypobaric hypoxia, which induces various physiological and molecular changes. Recent studies point toward interaction between circadian rhythms and the hypoxic response, yet their human relevance is lacking. Here, we examine the effect of different high altitudes in conjunction with time of day on human whole-blood transcriptome upon an expedition to the highest city in the world, La Rinconada, Peru, which is 5,100 m above sea level. We find that high altitude vastly affects the blood transcriptome and, unexpectedly, does not necessarily follow a monotonic response to altitude elevation. Importantly, we observe daily variance in gene expression, especially immune-related genes, which is largely altitude dependent. Moreover, using a digital cytometry approach, we estimate relative changes in abundance of different cell types and find that the response of several immune cell types is time- and altitude dependent. Taken together, our data provide evidence for interaction between the transcriptional response to hypoxia and the time of day in humans.
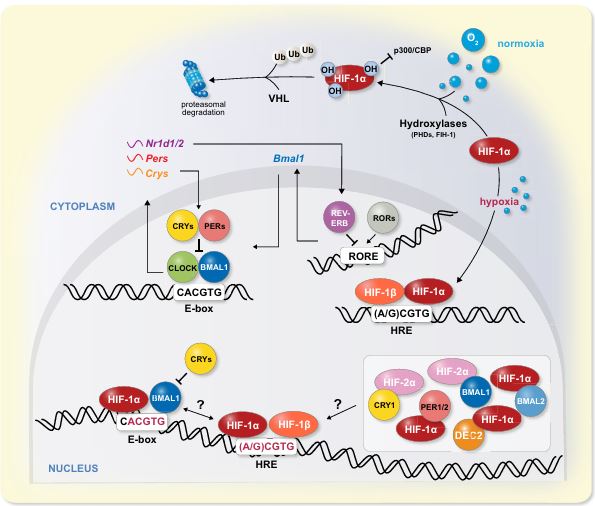
In mammals, physiology and metabolism are shaped both by immediate and anticipatory responses to environmental changes through the myriad of molecular mechanisms. Whilst the former is mostly mediated through different acute signalling pathways the latter is primarily orchestrated by the circadian clock. Oxygen is vital for life and as such mammals have evolved different mechanisms to cope with changes in oxygen levels. It is widely accepted that oxygen sensing through the HIF-1 signalling pathway is paramount for the acute response to changes in oxygen levels. Circadian clocks are molecular oscillators that control 24 hours rhythms in various aspects of physiology and behaviour. Evidence emerging in recent years points towards pervasive molecular and functional interactions between these two pathways on multiple levels. Daily oscillations in oxygen levels are circadian clock-controlled and can reset the clock through HIF-1. Furthermore, the circadian clock appears to modulate the hypoxic response. We review herein the literature related to the crosstalk between the circadian clockwork and the oxygen-signalling pathway in mammals at the molecular and physiological level both under normal and pathologic conditions.
Circadian clocks are self-sustained and cell-autonomous oscillators. They respond to various extracellular cues depending on the time-of-day and the signal intensity. Phase Transition Curves (PTCs) are instrumental in uncovering the full repertoire of responses to a given signal. However, the current methodologies for reconstructing PTCs are low-throughput, laborious, and resource- and time-consuming. We report here the development of an efficient and high throughput assay, dubbed Circadian Single-Cell Oscillators PTC Extraction (Circa-SCOPE) for generating high-resolution PTCs. This methodology relies on continuous monitoring of single-cell oscillations to reconstruct a full PTC from a single culture, upon a one-time intervention. Using Circa-SCOPE, we characterize the effects of various pharmacological and blood-borne resetting cues, at high temporal resolution and a wide concentration range. Thus, Circa-SCOPE is a powerful tool for comprehensive analysis and screening for circadian clocks resetting cues, and can be valuable for basic as well as translational research.
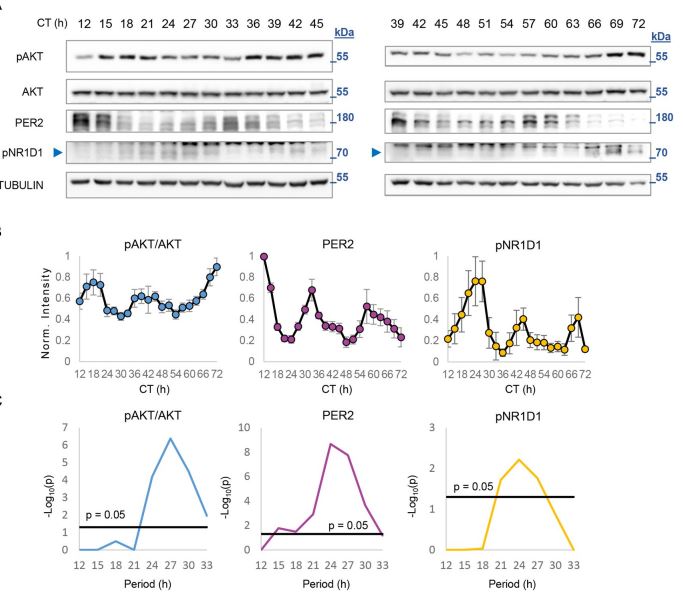
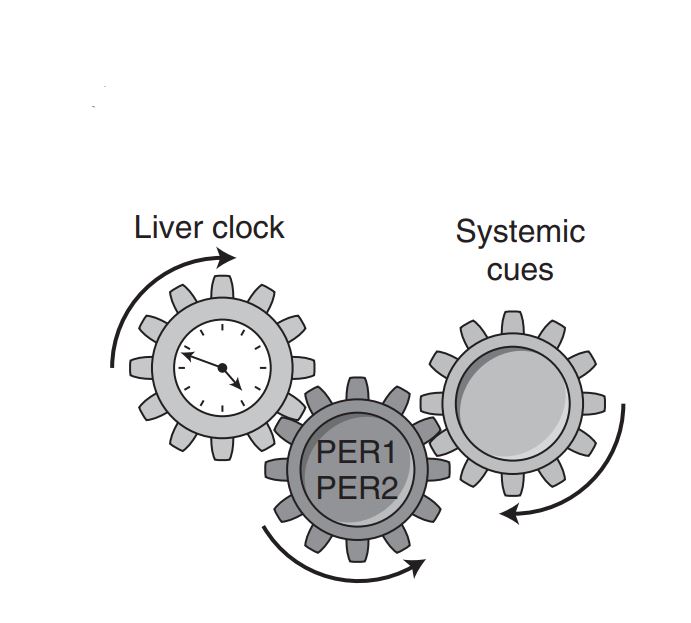
Rhythmicity of biological processes can be elicited either in response to environmental cycles or driven by endogenous oscillators. In mammals, the circadian clock drives about 24-hour rhythms of multitude metabolic and physiological processes in anticipation to environmental daily oscillations. Also at the intersection of environment and metabolism is the protein kinaseAKT. It conveys extracellular signals, primarily feeding-related signals, to regulate various key cellular functions. Previous studies in mice identified rhythmicity in AKT activation (pAKT) with elevated levels in the fed state. However, it is still unknown whether rhythmic AKT activation can be driven through intrinsic mechanisms. Here, we inspected temporal changes in pAKT levels both in cultured cells and animal models. In cultured cells, pAKT levels showed circadian oscillations similar to those observed in livers of wild-type mice under free-running conditions. Unexpectedly, in livers of Per1,2−/− but not of Bmal1−/− mice we detected ultradian (about 16 hours) oscillations of pAKT levels. Importantly, the liver transcriptome of Per1,2−/− mice also showed ultradian rhythms, corresponding to pAKT rhythmicity and consisting of AKT-related genes and regulators. Overall, our findings reveal ultradian rhythms in liver gene expression and AKT phosphorylation that emerge in the absence of environmental rhythms and Per1,2−/− genes.
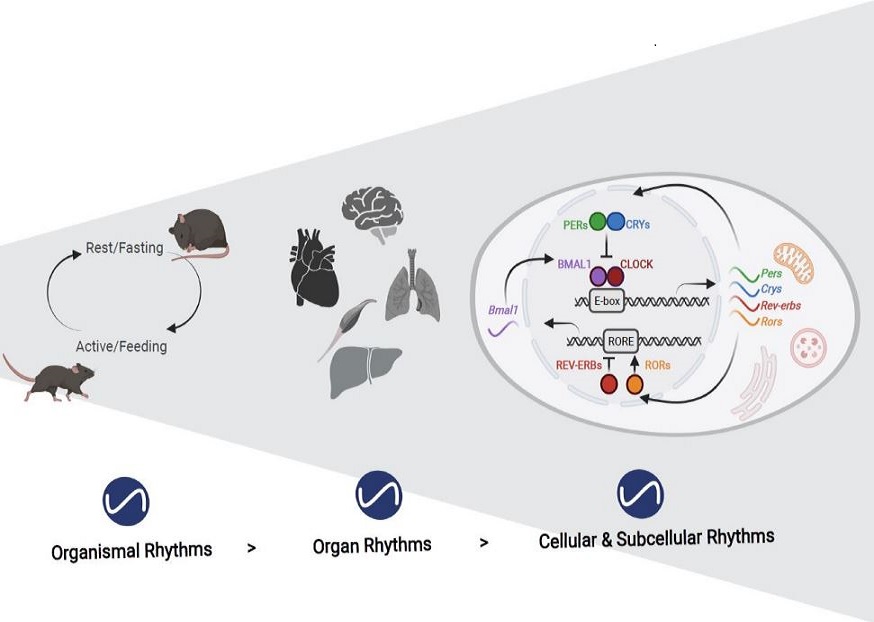
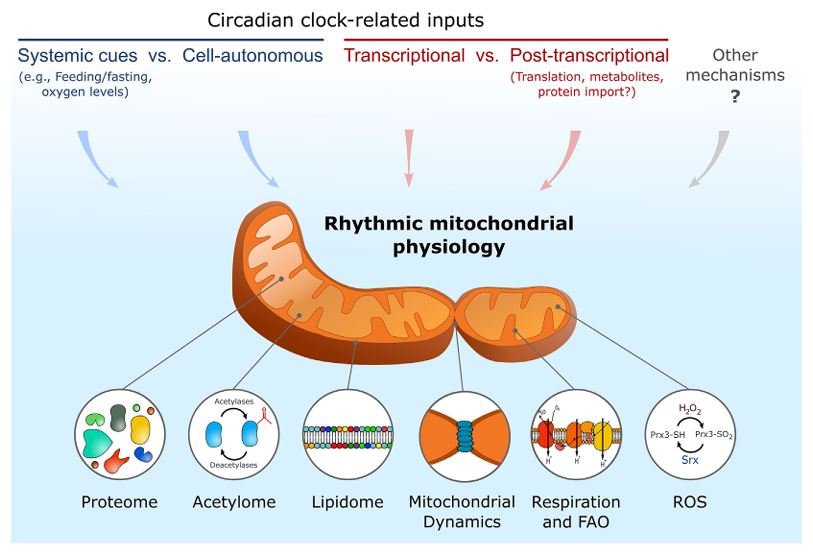
Circadian clocks have evolved in most lightsensitive organisms, from unicellular organisms to mammals. Consequently, a myriad of biological functions exhibits circadian rhythmicity, from behavior to physiology, through tissue and cellular functions to subcellular processes. Circadian rhythms in intracellular organelles are an emerging and exciting research arena. We summa-rize herein the current literature for rhythmicity in major intracellular organelles in mammals. These include changes in the morphology, content, and functions of different intracellular organelles. While these data highlight the presence of rhythmicity in these organelles, a gap remains in our knowledge regarding the underlying molecular mechanisms and their functional significance. Fi-nally, we discuss the importance and challenges faced by spatiotemporal studies on these organelles and speculate on the presence of oscillators in organelles and their potential mode of commu-nication. As circadian biology has been and continues to be studied throughout temporal and spatial axes, circadian organelles appear to be the next frontier.
Adamovich Y., Dandavate V., Ezagouri S., Manella G., Zwighaft Z., Sobel J., Kuperman Y., Golik M., Auerbach A., Itkin M., Malitsky S. & Asher G.
(2021)
Proceedings of the National Academy of Sciences - PNAS.
118,
35,
e210111511.
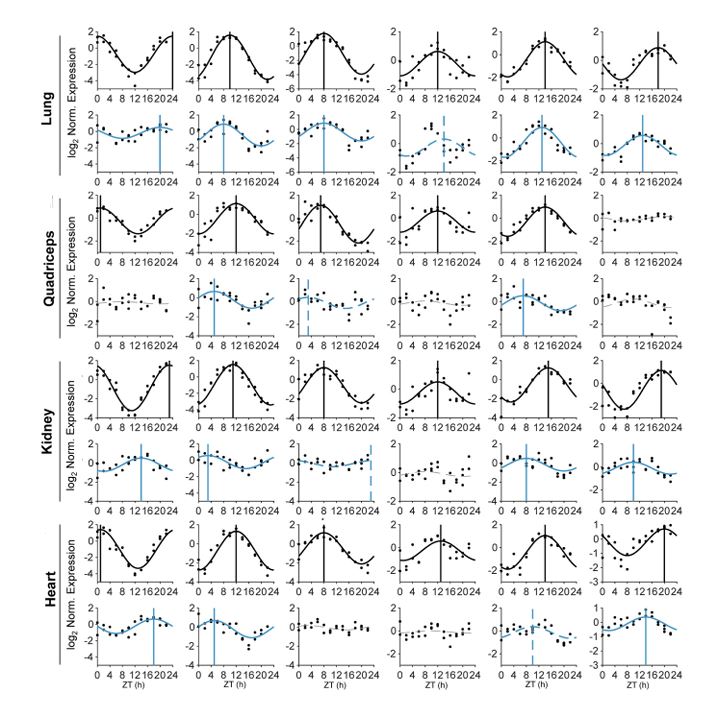
Exercise and circadian biology are closely intertwined with physiology and metabolism, yet the functional interaction between circadian clocks and exercise capacity is only partially characterized. Here, we tested different clock mutant mouse models to examine the effect of the circadian clock and clock proteins, namely PERIODs and BMAL1, on exercise capacity. We found that daytime variance in endurance exercise capacity is circadian clock controlled. Unlike wild-type mice, which outperform in the late compared with the early part of their active phase, PERIODs- and BMAL1-null mice do not show daytime variance in exercise capacity. It appears that BMAL1 impairs and PERIODs enhance exercise capacity in a daytime-dependent manner. An analysis of liver and muscle glycogen stores as well as muscle lipid utilization suggested that these daytime effects mostly relate to liver glycogen levels and correspond to the animals feeding behavior. Furthermore, given that exercise capacity responds to training, we tested the effect of training at different times of the day and found that training in the late compared with the early part of the active phase improves exercise performance. Overall, our findings suggest that clock proteins shape exercise capacity in a daytime-dependent manner through changes in liver glycogen levels, likely due to their effect on animals feeding behavior.
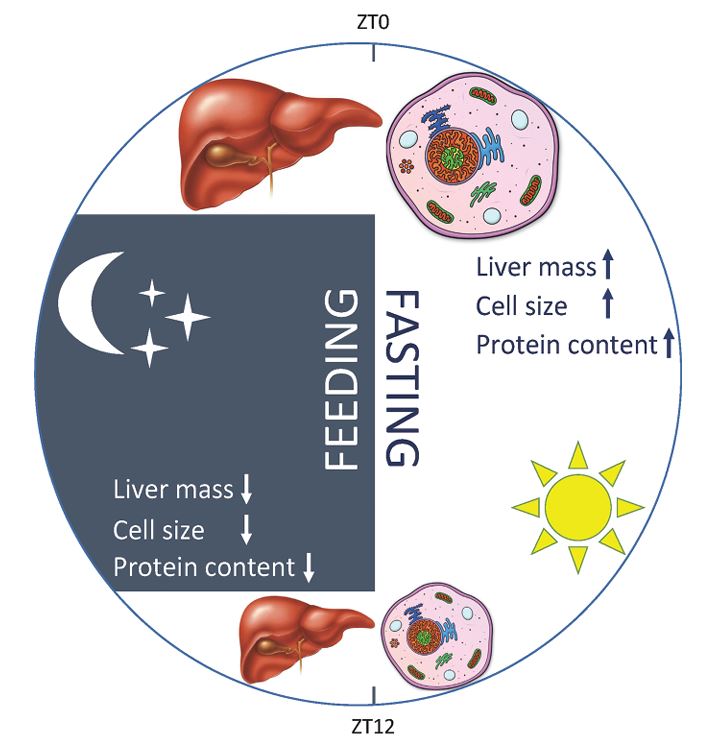
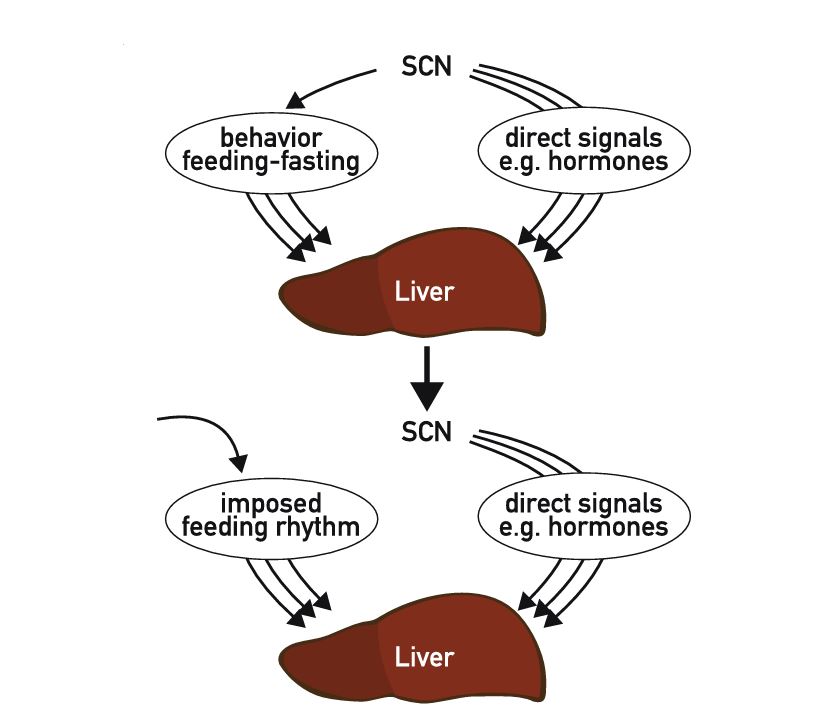
The mammalian circadian system consists of a central clock in the brain that synchronizes clocks in the peripheral tissues. Although the hierarchy between central and peripheral clocks is established, little is known regarding the specificity and functional organization of peripheral clocks. Here, we employ altered feeding paradigms in conjunction with liver-clock mutant mice to map disparities and interactions between peripheral rhythms. We find that peripheral clocks largely differ in their responses to feeding time. Disruption of the liver-clock, despite its prominent role in nutrient processing, does not affect the rhythmicity of clocks in other peripheral tissues. Yet, unexpectedly, liver-clock disruption strongly modulates the transcriptional rhythmicity of peripheral tissues, primarily on daytime feeding. Concomitantly, liver-clock mutant mice exhibit impaired glucose and lipid homeostasis, which are aggravated by daytime feeding. Overall, our findings suggest that, upon nutrient challenge, the liver-clock buffers the effect of feeding-related signals on rhythmicity of peripheral tissues, irrespective of their clocks.
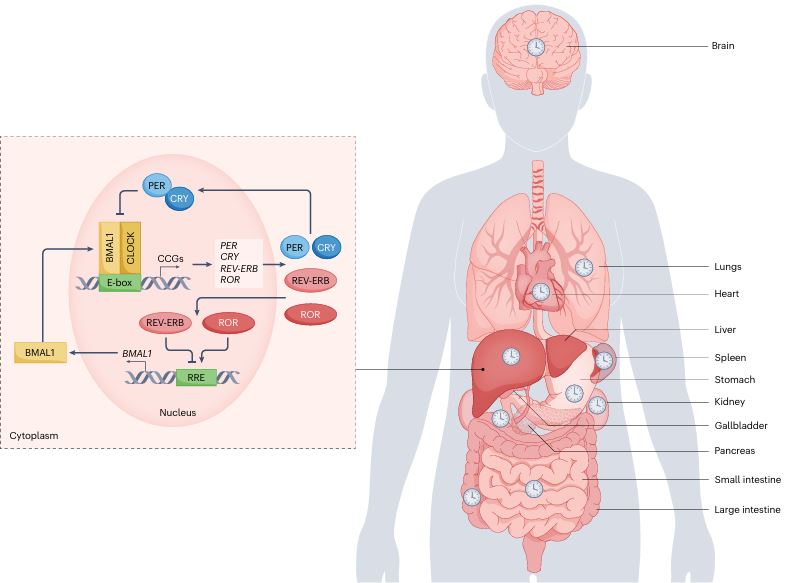
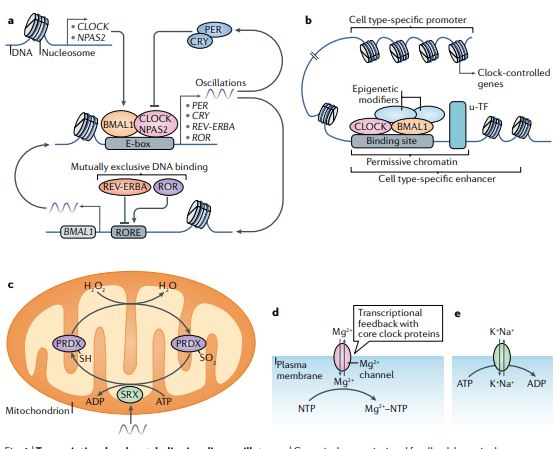
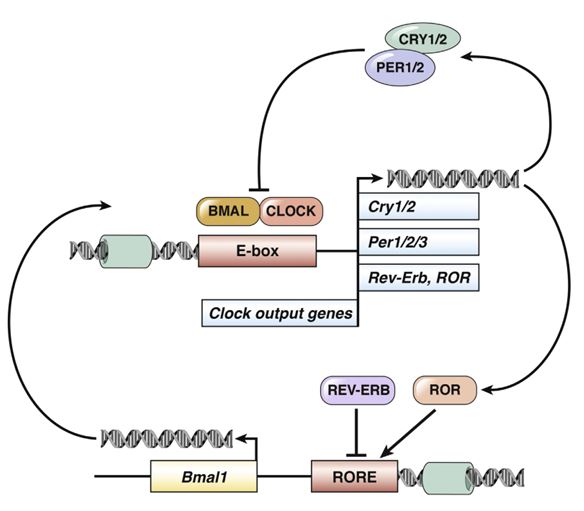
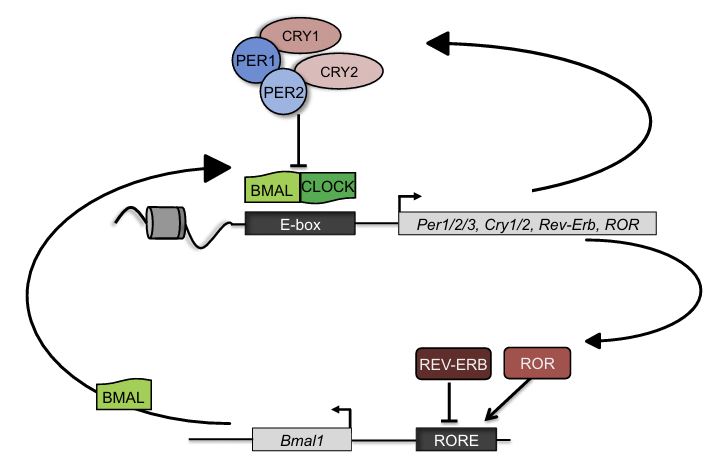
Circadian clocks oscillate over a period of ≈24 h in light-sensitive organisms and coordinate a wide variety of behavioral, physiological, and molecular functions with geophysical time. In mammals, clocks are present in virtually every cell of the body and function in a cell autonomous and self-sustained manner. The molecular clockwork relies on transcription-translation feedback loops, which generate self-sustained oscillations in the expression levels of the clock components (e.g. PERs, CRYs, CLOCK, BMAL1, NR1D1,2, RORs). These oscillations further control downstream processes through transcriptional and post-transcriptional regulation.
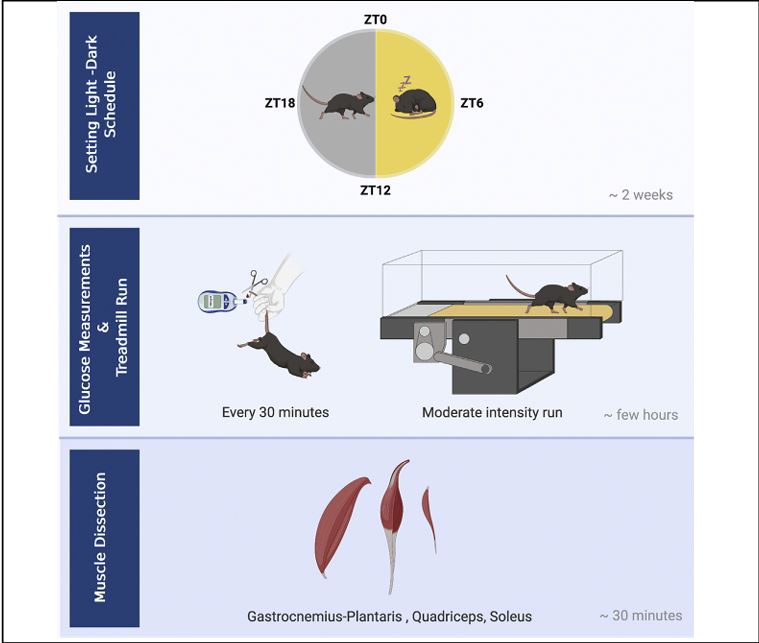
There is growing interest in medicine and sports in uncovering exercise modifiers that enhance or limit exercise capacity. Here, we detail a protocol for testing the daytime effect on running capacity in mice using a moderate intensity treadmill effort test. Instructions for dissecting soleus, gastrocnemius plantaris, and quadriceps muscles for further analysis are provided as well. This experimental setup is optimized for addressing questions regarding the involvement of daytime and circadian clocks in regulating exercise capacity. For complete details on the use and execution of this protocol, please refer to Ezagouri et al. (2019).
Lipidomics approaches provide quantitative characterization of hundreds of lipid species from biological samples. Recent studies highlight the value of these methods in studying circadian biology, and their potential goes far beyond studying lipid metabolism per se. For example, lipidomics analyses of subcellular compartments can be used to determine daily rhythmicity of different organelles and their intracellular dynamics. In this chapter we describe in detail the procedure for around the clock shotgun lipidomics, from sample preparation to bioinformatics analyses. Sample preparation includes biochemical fractionation of nuclei and mitochondria from mouse liver harvested throughout the day. Lipid content is determined and quantified, in unbiased manner and with wide coverage, using multidimensional mass spectrometry shotgun lipidomics (MDMS-SL). Circadian parameters are then determined with nonparametric statistical tests. Overall, the approach described herein is applicable for various animal models, tissues, and organelles, and is expected to yield new insight on various aspects of circadian biology and lipid metabolism.
Manella G., Aviram R., Bolshette N., Muvkadi S., Golik M., Smith D. F. & Asher G.
(2020)
Proceedings of the National Academy of Sciences of the United States of America.
117,
1,
p. 779-786
The occurrence and sequelae of disorders that lead to hypoxic spells such as asthma, chronic obstructive pulmonary disease, and obstructive sleep apnea (OSA) exhibit daily variance. This prompted us to examine the interaction between the hypoxic response and the circadian clock in vivo. We found that the global transcriptional response to acute hypoxia is tissue-specific and time-of-day-dependent. In particular, clock components differentially responded at the transcriptional and posttranscriptional level, and these responses depended on an intact circadian clock. Importantly, exposure to hypoxia phase-shifted clocks in a tissue-dependent manner led to intertissue circadian clock misalignment. This differential response relied on the intrinsic properties of each tissue and could be recapitulated ex vivo. Notably, circadian misalignment was also elicited by intermittent hypoxia, a widely used model for OSA. Given that phase coherence between circadian clocks is considered favorable, we propose that hypoxia leads to circadian misalignment, contributing to the pathophysiology of OSA and potentially other diseases that involve hypoxia.
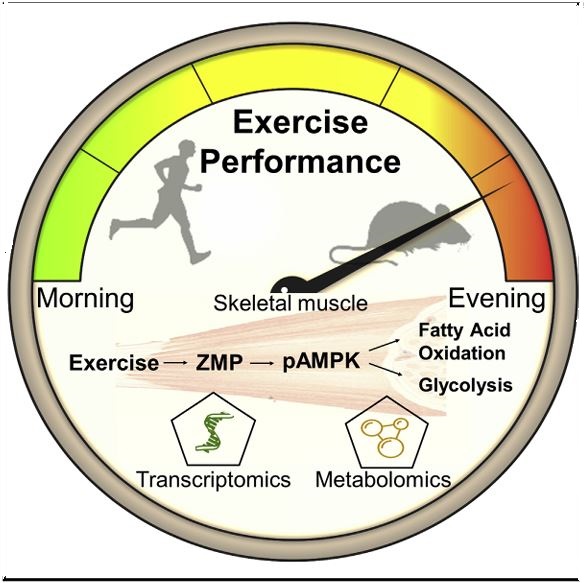
Physical performance relies on the concerted action of myriad responses, many of which are under circadian clock control. Little is known, however, regarding the time-dependent effect on exercise performance at the molecular level. We found that both mice and humans exhibit daytime variance in exercise capacity between the early and late part of their active phase. The daytime variance in mice was dependent on exercise intensity and relied on the circadian clock proteins PER1/2. High-throughput gene expression and metabolic profiling of skeletal muscle revealed metabolic pathways that are differently activated upon exercise in a daytime-dependent manner. Remarkably, we discovered that ZMP, an endogenous AMPK activator, is induced by exercise in a time-dependent manner to regulate key steps in glycolytic and fatty acid oxidation pathways and potentially enhance exercise capacity. Overall, we propose that time of day is a major modifier of exercise capacity and associated metabolic pathways.
Adamovich Y., Ladeuix B., Sobel J., Manella G., Neufeld-Cohen A., Assadi M. H., Golik M., Kuperman Y., Tarasiuk A., Koeners M. P. & Asher G.
(2019)
Cell Metabolism.
29,
5,
p. 1092-1103.e3
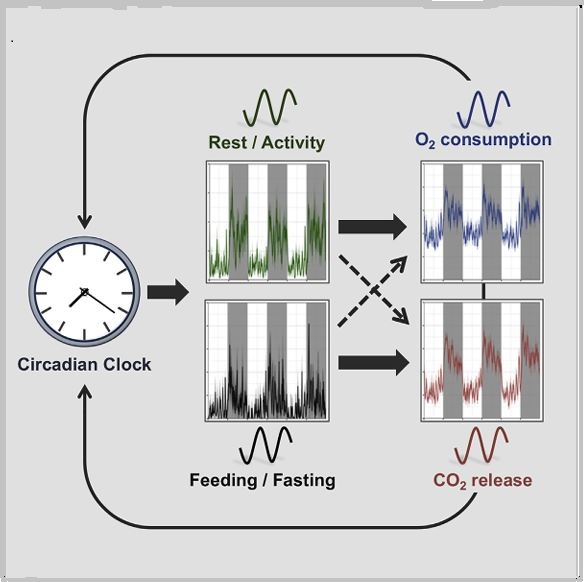
Daily rhythms in animal physiology are driven by endogenous circadian clocks in part through rest-activity and feeding-fasting cycles. Here, we examined principles that govern daily respiration. We monitored oxygen consumption and carbon dioxide release, as well as tissue oxygenation in freely moving animals to specifically dissect the role of circadian clocks and feeding time on daily respiration. We found that daily rhythms in oxygen and carbon dioxide are clock controlled and that time-restricted feeding restores their rhythmicity in clock-deficient mice. Remarkably, day-time feeding dissociated oxygen rhythms from carbon dioxide oscillations, whereby oxygen followed activity, and carbon dioxide was shifted and aligned with food intake. In addition, changes in carbon dioxide levels altered clock gene expression and phase shifted the clock. Collectively, our findings indicate that oxygen and carbon dioxide rhythms are clock controlled and feeding regulated and support a potential role for carbon dioxide in phase resetting peripheral clocks upon feeding. Adamovich et al. show that the daily regulation of oxygen consumption and carbon dioxide release is regulated by the circadian clock. Time-restricted feeding restores oxygen and carbon dioxide rhythms in clock mutants. Their findings also support a potential role for carbon dioxide in phase resetting of peripheral clocks upon feeding
Humans, like all mammals, partition their daily behaviour into activity (wakefulness) and rest (sleep) phases that differ largely in their metabolic requirements. The circadian clock evolved as an autonomous timekeeping system that aligns behavioural patterns with the solar day and supports the body functions by anticipating and coordinating the required metabolic programmes. The key component of this synchronization is a master clock in the brain, which responds to light-darkness cues from the environment. However, to achieve circadian control of the entire organism, each cell of the body is equipped with its own circadian oscillator that is controlled by the master clock and confers rhythmicity to individual cells and organs through the control of rate-limiting steps of metabolic programmes. Importantly, metabolic regulation is not a mere output function of the circadian system, but nutrient, energy and redox levels signal back to cellular clocks in order to reinforce circadian rhythmicity and to adapt physiology to temporal tissue-specific needs. Thus, multiple systemic and molecular mechanisms exist that connect the circadian clock with metabolism at all levels, from cellular organelles to the whole organism, and deregulation of this circadian-metabolic crosstalk can lead to various pathologies.
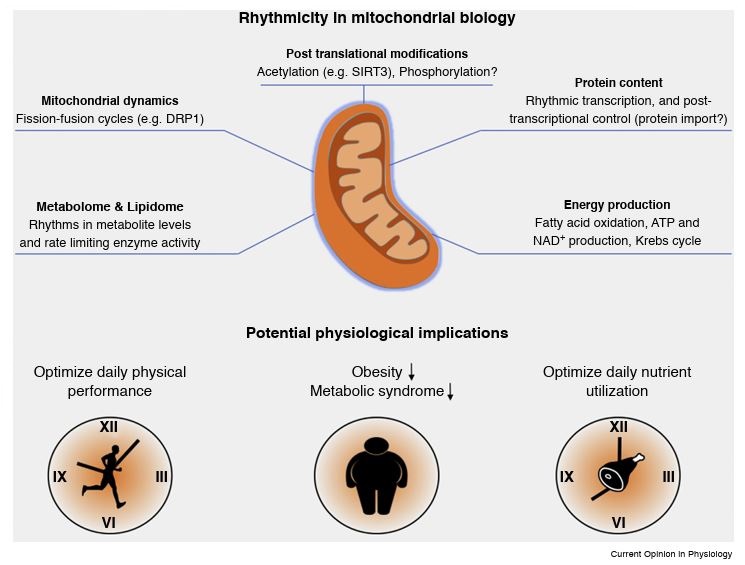
Physiology and behavior in mammals is predominantly under circadian clock control. Circadian clocks are present in nearly every cell of the body and oscillate with a frequency of about 24 h in a self-sustained and cell autonomous manner. These clocks not only play a principal role in metabolic control but concurrently respond to a wide variety of metabolic cues. While various aspects of circadian regulation of metabolic functions have been extensively studied, our knowledge regarding circadian mitochondrial biology is just emerging. We review herein the current literature addressing circadian mitochondrial biology: from diurnal changes in mitochondrial make-up, through dynamics and functions. We will discuss as well potential mechanisms that are implicated in circadian control of mitochondrial biology in mammals.
Hughes M. E., Abruzzi K. C., Allada R., Anafi R., Arpat A. B., Asher G., Baldi P., de Bekker C., Bell-Pedersen D., Blau J., Brown S., Ceriani M. F., Chen Z., Chiu J. C., Cox J., Crowell A. M., DeBruyne J. P., Dijk D. J., DiTacchio L., Doyle F. J., Duffield G. E., Dunlap J. C., Eckel-Mahan K., Esser K. A., FitzGerald G. A., Forger D. B., Francey L. J., Fu Y. H., Gachon F., Gatfield D., de Goede P., Golden S. S., Green C., Harer J., Harmer S., Haspel J., Hastings M. H., Herzel H., Herzog E. D., Hoffmann C., Hong C., Hughey J. J., Hurley J. M., de la Iglesia H. O., Johnson C., Kay S. A., Koike N., Kornacker K., Kramer A., Lamia K., Leise T., Lewis S. A., Li J., Li X., Liu A. C., Loros J. J., Martino T. A., Menet J. S., Merrow M., Millar A. J., Mockler T., Naef F., Nagoshi E., Nitabach M. N., Olmedo M., Nusinow D. A., Ptáček L. J., Rand D., Reddy A. B., Robles M. S., Roenneberg T., Rosbash M., Ruben M. D., Rund S. S., Sancar A., Sassone-Corsi P., Sehgal A., Sherrill-Mix S., Skene D. J., Storch K. F., Takahashi J. S., Ueda H. R., Wang H., Weitz C., Westermark P. O., Wijnen H., Xu Y., Wu G., Yoo S. H., Young M., Zhang E. E., Zielinski T. & Hogenesch J. B.
(2017)
Journal of Biological Rhythms.
32,
5,
p. 380-393
Genome biology approaches have made enormous contributions to our understanding of biological rhythms, particularly in identifying outputs of the clock, including RNAs, proteins, and metabolites, whose abundance oscillates throughout the day. These methods hold significant promise for future discovery, particularly when combined with computational modeling. However, genome-scale experiments are costly and laborious, yielding \u201cbig data\u201d that are conceptually and statistically difficult to analyze. There is no obvious consensus regarding design or analysis. Here we discuss the relevant technical considerations to generate reproducible, statistically sound, and broadly useful genome-scale data. Rather than suggest a set of rigid rules, we aim to codify principles by which investigators, reviewers, and readers of the primary literature can evaluate the suitability of different experimental designs for measuring different aspects of biological rhythms. We introduce CircaInSilico, a web-based application for generating synthetic genome biology data to benchmark statistical methods for studying biological rhythms. Finally, we discuss several unmet analytical needs, including applications to clinical medicine, and suggest productive avenues to address them.
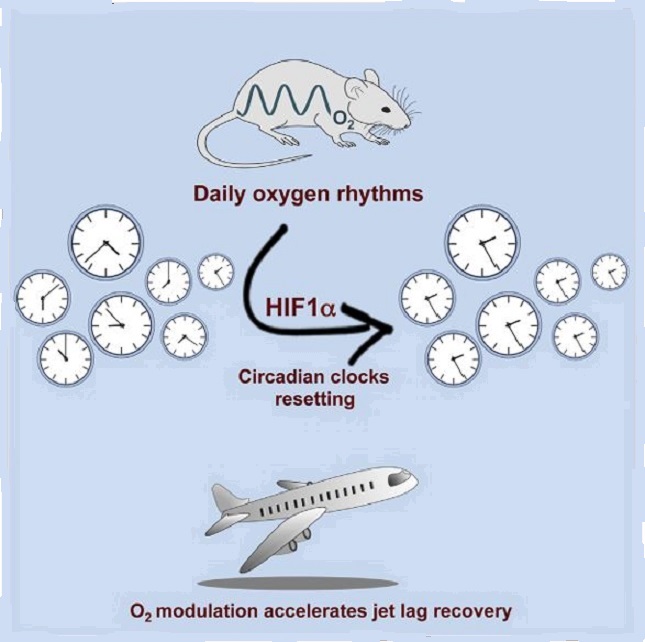
The mammalian circadian system consists of a master clock in the brain that synchronizes subsidiary oscillators in peripheral tissues. The master clock maintains phase coherence in peripheral cells through systemic cues such as feeding-fasting and temperature cycles. Here, we examined the role of oxygen as a resetting cue for circadian clocks. We continuously measured oxygen levels in living animals and detected daily rhythms in tissue oxygenation. Oxygen cycles, within the physiological range, were sufficient to synchronize cellular clocks in a HIF1α-dependent manner. Furthermore, several clock genes responded to changes in oxygen levels through HIF1α. Finally, we found that a moderate reduction in oxygen levels for a short period accelerates the adaptation of wild-type but not of HIF1α-deficient mice to the new time in a jet lag protocol. We conclude that oxygen, via HIF1α activation, is a resetting cue for circadian clocks and propose oxygen modulation as therapy for jet lag.
Circadian clocks orchestrate the daily changes in physiology and behavior of light-sensitive organisms. These clocks measure about 24 h and tick in a self-sustained and cell-autonomous manner. Mounting evidence points toward a tight intertwining between circadian clocks and metabolism. Although various aspects of circadian control of metabolic functions have been extensively studied, our knowledge regarding circadian mitochondrial function is rudimentary. In this review, we will survey the current literature related to the circadian nature of mitochondrial biology: from mitochondrial omics studies (e.g., proteome, acetylome, and lipidome), through dissection of mitochondrial morphology, to analyses of mitochondrial processes such as nutrient utilization and respiration. We will describe potential mechanisms that are implicated in circadian regulation of mitochondrial functions in mammals and discuss the possibility of a mitochondrial-autonomous oscillator.
Aviram R., Manella G., Kopelman N., Neufeld - Cohen -. C. A., Zwighaft Z., Elimelech M., Adamovich Y., Golik M., Wang C., Han X. & Asher G.
(2016)
Molecular Cell.
62,
4,
p. 636-648
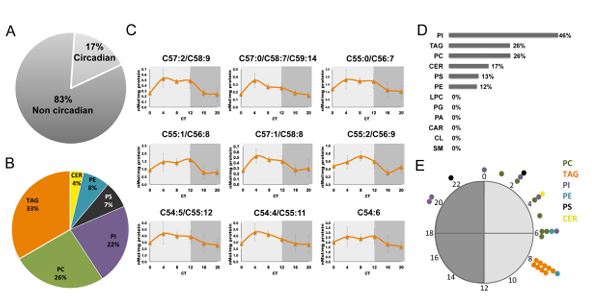
Cells have evolved mechanisms to handle incompatible processes through temporal organization by circadian clocks and by spatial compartmentalization within organelles defined by lipid bilayers. Recent advances in lipidomics have led to identification of plentiful lipid species, yet our knowledge regarding their spatiotemporal organization is lagging behind. In this study, we quantitatively characterized the nuclear and mitochondrial lipidome in mouse liver throughout the day, upon different feeding regimens, and in clock-disrupted mice. Our analyses revealed potential connections between lipid species within and between lipid classes. Remarkably, we uncovered diurnal oscillations in lipid accumulation in the nucleus and mitochondria. These oscillations exhibited opposite phases and readily responded to feeding time. Furthermore, we found that the circadian clock coordinates the phase relation between the organelles. In summary, our study provides temporal and spatial depiction of lipid organization and reveals the presence and coordination of diurnal rhythmicity in intracellular organelles. Aviram et al. performed lipidomics analyses of organelles from mouse liver to explore temporal and spatial principles in lipid organization. They find that the lipid composition of the nucleus and mitochondria oscillates daily with distinct and opposite phases. These oscillations respond to feeding time and are coordinated by the circadian clock.
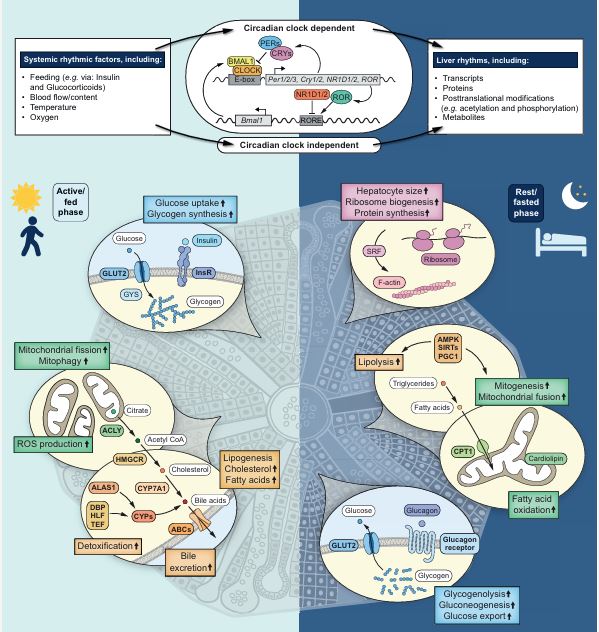
Our understanding of the molecular underpinnings of the mammalian circadian oscillator and its tight connection to physiology has progressed tremendously during the past decades. The liver is considered the prototypic experimental model tissue for circadian research in peripheral organs. Studies on liver clocks have been highly productive and yielded information about widely different aspects of circadian biology. The liver, as one of the largest organs in the body, has often been used for the identification of core clock and auxiliary clock components, for example, by biochemical purifications. Because the liver is also a major metabolic hub, studies addressing the interplay between circadian clocks and metabolism have been insightful. In addition, the use of liver-specific loss-of-function models for clock components highlighted not only specific physiological roles of the hepatic clock but also its interplay with systemic cues and oscillators in other organs. Recently, technological advances in omics approaches have been successfully applied on the liver, providing a comprehensive depiction of pervasive circadian control of gene expression and protein and metabolite accumulation. In this review, we chose to illuminate specific examples that demonstrate how different experimental approaches - namely, biochemical, metabolic, genetic, and omics methodologies - have advanced our knowledge regarding circadian liver biology and chronobiology in general.
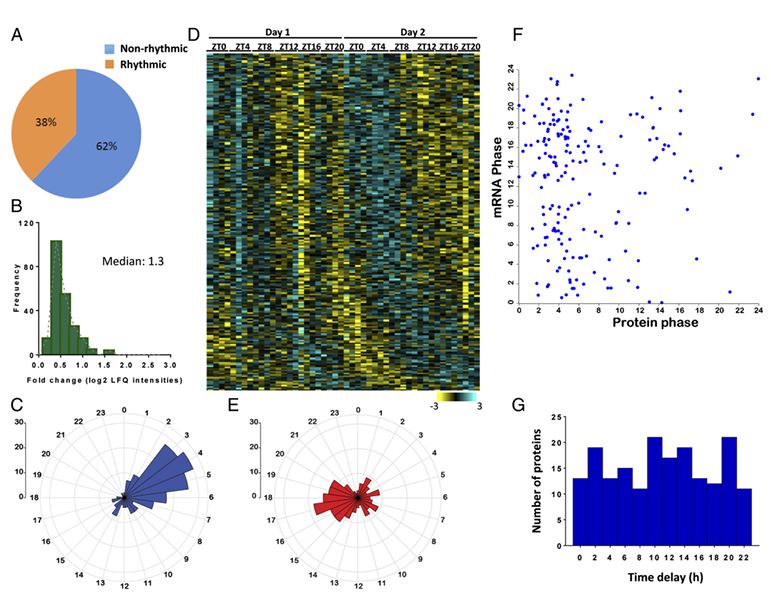
Neufeld - Cohen A., Robles M., Aviram R., Manella G., Adamovich Y., Ladeuix B., Nir D., Rousso Noori N. L., Kuperman Y., Golik M., Mann M. & Asher G.
(2016)
Proceedings of the National Academy of Sciences of the United States of America.
113,
12,
p. E1673-E1682
Mitochondria are major suppliers of cellular energy through nutrients oxidation. Little is known about the mechanisms that enable mitochondria to cope with changes in nutrient supply and energy demand that naturally occur throughout the day. To address this question, we applied MS-based quantitative proteomics on isolated mitochondria from mice killed throughout the day and identified extensive oscillations in the mitochondrial proteome. Remarkably, the majority of cycling mitochondrial proteins peaked during the early light phase. We found that rate-limiting mitochondrial enzymes that process lipids and carbohydrates accumulate in a diurnal manner and are dependent on the clock proteins PER1/2. In this conjuncture, we uncovered daily oscillations in mitochondrial respiration that peak during different times of the day in response to different nutrients. Notably, the diurnal regulation of mitochondrial respiration was blunted in mice lacking PER1/2 or on a high-fat diet. We propose that PERIOD proteins optimize mitochondrial metabolism to daily changes in energy supply/demand and thereby, serve as a rheostat for mitochondrial nutrient utilization.
The circadian clock is an endogenous biological timekeeping system that synchronizes physiology and behavior to day/night cycles. A wide variety of processes throughout the entire gastrointestinal tract and notably the liver appear to be under circadian control. These include various metabolic functions such as nutrient uptake, processing, and detoxification, which align organ function to cycle with nutrient supply and demand. Remarkably, genetic or environmental disruption of the circadian clock can cause metabolic diseases or exacerbate pathological states. In addition, modern lifestyles force more and more people worldwide into asynchrony between the external time and their circadian clock, resulting in a constant state of social jetlag. Recent evidence indicates that interactions between altered energy metabolism and disruptions in the circadian clock create a downward spiral that can lead to diabetes and other metabolic diseases. In this review, we provide an overview of rhythmic processes in the liver and highlight the functions of circadian clock genes under physiological and pathological conditions; we focus on their roles in regulation of hepatic glucose as well as lipid and bile acid metabolism and detoxification and their potential effects on the development of fatty liver and nonalcoholic steatohepatitis.
Zwighaft Z., Aviram R., Shalev M., Rousso Noori N. L., Kraut-Cohen J., Golik M., Brandis A., Reinke H., Aharoni A., Kahana C. & Asher G.
(2015)
Cell Metabolism.
22,
5,
p. 874-885
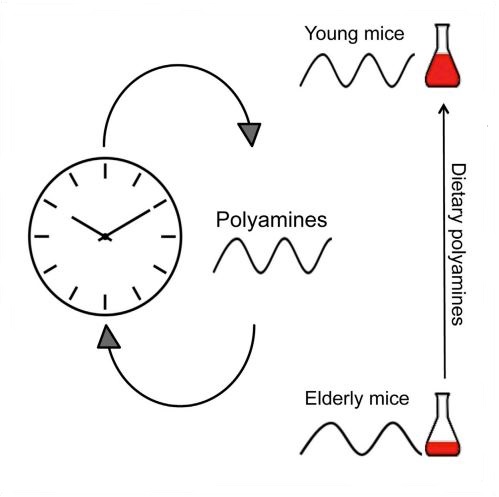
Polyamines are essential polycations present in all living cells. Polyamine levels are maintained from the diet and de novo synthesis, and their decline with age is associated with various pathologies. Here we show that polyamine levels oscillate in a daily manner. Both clock- and feeding-dependent mechanisms regulate the daily accumulation of key enzymes in polyamine biosynthesis through rhythmic binding of BMAL1:CLOCK to conserved DNA elements. In turn, polyamines control the circadian period in cultured cells and animals by regulating the interaction between the core clock repressors PER2 and CRY1. Importantly, we found that the decline in polyamine levels with age in mice is associated with a longer circadian period that can be reversed upon polyamine supplementation in the diet. Our findings suggest a crosstalk between circadian clocks and polyamine biosynthesis and open new possibilities for nutritional interventions against the decay in clock's function with age.
Lipids play vital roles in a wide variety of cellular functions. They act as structural components in cell membranes, serve as a major form of energy storage, and function as key signaling molecules. Mounting evidence points towards a tight interplay between lipids and circadian clocks. In mammals, circadian clocks regulate the daily physiology and metabolism, and disruption of circadian rhythmicity is associated with altered lipid homeostasis and pathologies such as fatty liver and obesity. Concomitantly, emerging evidence suggest that lipids are embedded within the core clock circuitry and participate in circadian control. Recent advances in lipidomics methodologies and their application in chronobiology studies have shed new light on the cross talk between circadian clocks and lipid homeostasis. We review herein the latest literature related to the involvement of lipids in circadian clock's function and highlight the contribution of circadian lipidomics studies to our understanding of circadian rhythmicity and lipid homeostasis. This article is part of a Special Issue entitled Brain Lipids.
The circadian clock, a highly specialized, hierarchical network of biological pacemakers, directs and maintains proper rhythms in endocrine and metabolic pathways required for organism homeostasis. The clock adapts to environmental changes, specifically daily light-dark cycles, as well as rhythmic food intake. Nutritional challenges reprogram the clock, while time-specific food intake has been shown to have profound consequences on physiology. Importantly, a critical role in the clock-nutrition interplay appears to be played by the microbiota. The circadian clock appears to operate as a critical interface between nutrition and homeostasis, calling for more attention on the beneficial effects of chrono-nutrition.
The circadian core clock circuitry relies on interlocked transcription-translation feedback loops that largely count on multiple protein interactions. The molecularmechanisms implicated in the assembly of these protein complexes are relatively unknown. Our bioinformatics analysis of short linear motifs, implicated in protein interactions, reveals an enrichment of the Pro-X-Asp-Leu-Ser (PXDLS) motif within circadian transcripts. We show that the PXDLS motif can bind to BMAL1/CLOCK and disrupt circadian oscillations in a cell-autonomous manner. Remarkably, the motif is evolutionary conserved in the core clock protein REV-ERBα, and additional proteins implicated in the clock's function (NRIP1, CBP). In this conjuncture, we uncover a novel cross talk between the two principal core clock feedback loops and show that BMAL/CLOCK and REV-ERBα interact and that the PXDLS motif of REV-ERBα participates in their binding. Furthermore, we demonstrate that the PXDLS motifs of NRIP1 and CBP are involved in circadian rhythmicity. Our findings suggest that the PXDLS motif plays an important role in circadian rhythmicity through regulation of protein interactions within the clock circuitry and that short linear motifs can be employed to modulate circadian oscillations.
Adamovich Y., Rousso-Noori L., Zwighaft Z., Neufeld-Cohen A., Golik M., Kraut-Cohen J., Wang M., Han X. & Asher G.
(2014)
Cell Metabolism.
19,
2,
p. 319-330
Circadian clocks play a major role in orchestrating daily physiology, and their disruption can evoke metabolic diseases such as fatty liver and obesity. To study the role of circadian clocks in lipid homeostasis, we performed an extensive lipidomic analysis of liver tissues from wild-type and clock-disrupted mice either fed ad libitum or night fed. To our surprise, a similar fraction of lipids (∼17%) oscillated in both mouse strains, most notably triglycerides, but with completely different phases. Moreover, several master lipid regulators (e.g., PPARα) and enzymes involved in triglyceride metabolism retained their circadian expression in clock-disrupted mice. Nighttime restricted feeding shifted the phase of triglyceride accumulation and resulted in ∼50% decrease in hepatic triglyceride levels in wild-type mice. Our findings suggest that circadian clocks and feeding time dictate the phase and levels of hepatic triglyceride accumulation; however, oscillations in triglycerides can persist in the absence of a functional clock.
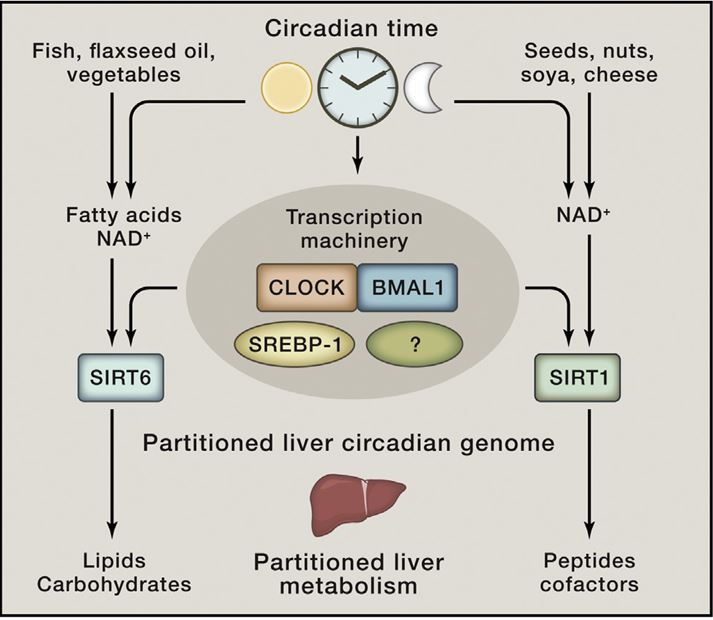
In mammals, most metabolic processes are influenced by biological clocks and feeding rhythms. The mechanisms that couple metabolism to circadian oscillators are just emerging. NAD-dependent enzymes (e.g., Sirtuins and poly[ADP-ribose] polymerases), redox- and/or temperature-dependent transcription factors (e.g., CLOCK, NPAS2, and HSF1), nutrient-sensing transcriptional regulatory proteins (e.g., CREB-CBP-CRCT2, FOXO-p300, nuclear receptors, PGC-1, and SP1 family members) and protein kinases (e.g., AMPK), are plausible candidates for conveying a cell's metabolic state to the core clock circuitry. The intertwining between these acute regulators and circadian clock components is so tight that the discrimination between metabolic and circadian oscillations may be somewhat arbitrary.
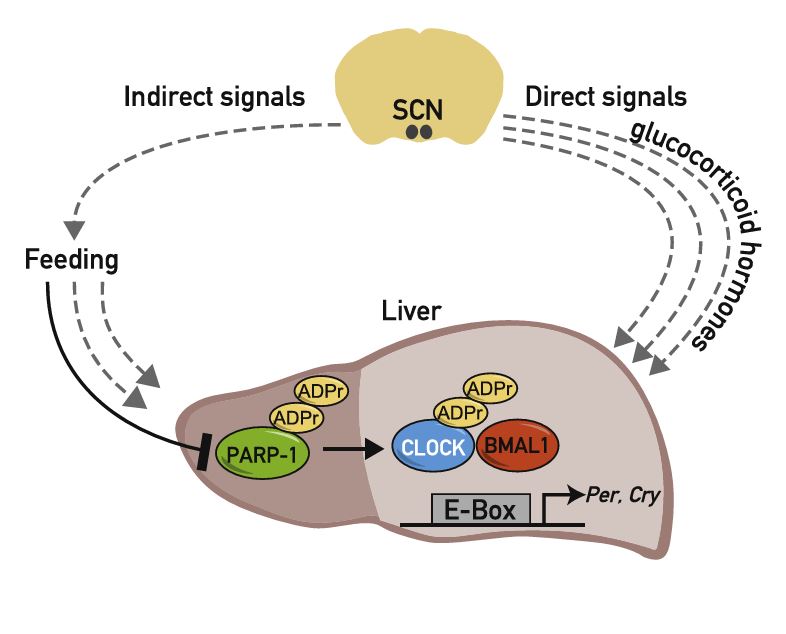
Circadian clocks in peripheral organs are tightly coupled to cellular metabolism and are readily entrained by feeding-fasting cycles. However, the molecular mechanisms involved are largely unknown. Here we show that in liver the activity of PARP-1, an NAD+-dependent ADP-ribosyltransferase, oscillates in a daily manner and is regulated by feeding. We provide biochemical evidence that PARP-1 binds and poly(ADP-ribosyl)ates CLOCK at the beginning of the light phase. The loss of PARP-1 enhances the binding of CLOCK-BMAL1 to DNA and leads to a phase-shift of the interaction of CLOCK-BMAL1 with PER and CRY repressor proteins. As a consequence, CLOCK-BMAL1-dependent gene expression is altered in PARP-1-deficient mice, in particular in response to changes in feeding times. Our results show that Parp-1 knockout mice exhibit impaired food entrainment of peripheral circadian clocks and support a role for PARP-1 in connecting feeding with the mammalian timing system. PaperFlick:
Most physiological processes are subject to daily-oscillations that are driven by an endogenous circadian clock. These include rest-activity cycles, cardiovascular functions such as heart rate and blood pressure, the production and secretion of hormones, renal plasma flow and urine production, in addition to metabolic functions of organs associated with the gastrointestinal tract (for review and references, see [1-4]). Since most metabolic functions oscillate in a daily manner, the liver is an organ for which circadian timing is particularly obvious. Thus, genome-wide transcriptome profiling studies have revealed that depending on the stringency of algorithms used for the extraction of oscillating transcripts between 2 and 10% of all liver mRNAs accumulate in a rhythmic fashion [5-10]. The majority of these transcripts encode enzymes and regulators involved in the metabolism of fatty acids, cholesterol, bile acids, carbohydrates, and xenobiotics. Several signaling pathways relevant for hepatic clock outputs (e.g., signaling through PPARs, CAR, LXR, and FXR) are elaborated in previous chapters of this issue. In this chapter, we shall thus focus on putative signaling pathways related to input pathways into the liver clock. Specifically, we will discuss current views and hypothesis on how the master pacemaker in the brain's suprachiasmatic nucleus (SCN) synchronizes peripheral clocks, in particular those operative in liver. We will also present some findings made with cultured fibroblasts, since these cells have served as a model system in most in vitro studies. Some of the signaling routes outlined below remain speculative, and their detailed analysis requires additional investigations.
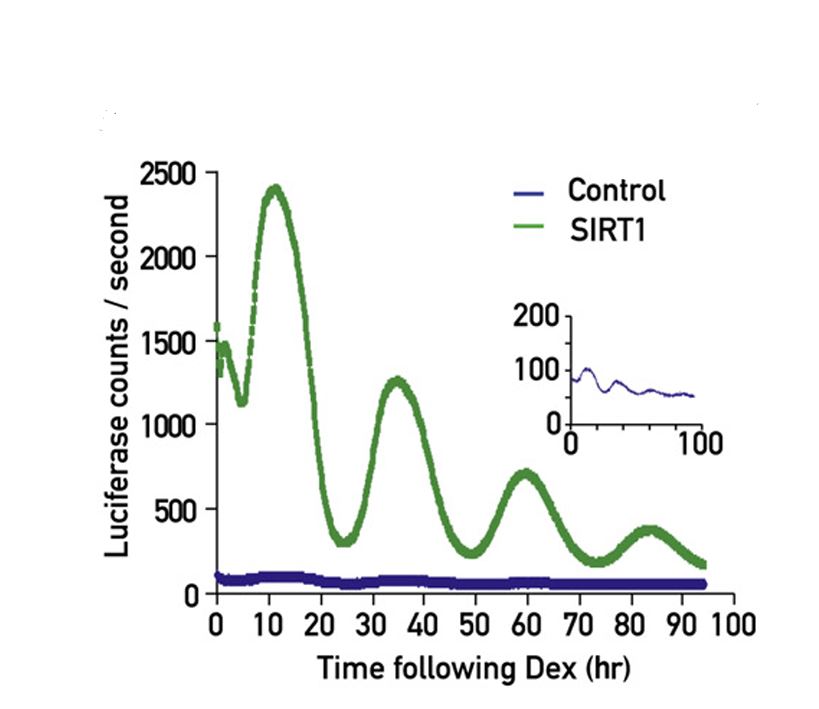
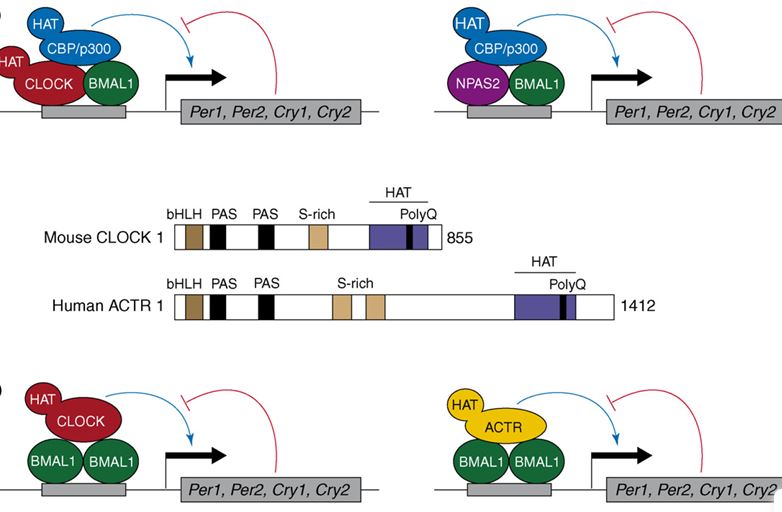
The mammalian circadian timing system is composed of a central pacemaker in the suprachiasmatic nucleus of the brain that synchronizes countless subsidiary oscillators in peripheral tissues. The rhythm-generating mechanism is thought to rely on a feedback loop involving positively and negatively acting transcription factors. BMAL1 and CLOCK activate the expression of Period (Per) and Cryptochrome (Cry) genes, and once PER and CRY proteins accumulate to a critical level they form complexes with BMAL1-CLOCK heterodimers and thereby repress the transcription of their own genes. Here, we show that SIRT1, an NAD+-dependent protein deacetylase, is required for high-magnitude circadian transcription of several core clock genes, including Bmal1, Rorγ, Per2, and Cry1. SIRT1 binds CLOCK-BMAL1 in a circadian manner and promotes the deacetylation and degradation of PER2. Given the NAD+ dependence of SIRT1 deacetylase activity, it is likely that SIRT1 connects cellular metabolism to the circadian core clockwork circuitry.
Mammalian physiology is governed by a complex circadian timing system that involves interacting positive and negative transcriptional feedback loops. A key role in this feedback loop was attributed to the PAS domain helix-loop-helix protein CLOCK, on the basis of a dominant-negative mutation in this transcription factor. However, recent experiments by Reppert and coworkers with Clock knockout mice suggest that CLOCK is dispensable for rhythmic gene expression and behavior, presumably because other proteins can substitute for CLOCK in these animals.
Tsvetkov P., Asher G., Reiss V., Shaul Y., Sachs L. & Lotem J.
(2005)
Proceedings of the National Academy of Sciences of the United States of America.
102,
15,
p. 5535-5540
NAD(P)H:quinone oxidoreductase 1 (NQO1) regulates the stability of the tumor suppressor WT p53. NQO1 binds and stabilizes WT p53, whereas NQO1 inhibitors including dicoumarol and various other coumarins and flavones induce ubiquitin-independent proteasomal p53 degradation and thus inhibit p53-induced apoptosis. Here, we show that curcumin, a natural phenolic compound found in the spice turmeric, induced ubiquitin-independent degradation of WT p53 and inhibited p53-induced apoptosis in normal thymocytes and myeloid leukemic cells. Like dicoumarol, curcumin inhibited the activity of recombinant NQO1 in vitro, inhibited the activity of endogenous cellular NQO1 in vivo, and dissociated NQO1-WT p53 complexes. Neither dicoumarol nor curcumin dissociated the complexes of NQO1 and the human cancer hot-spot p53 R273H mutant and therefore did not induce degradation of this mutant. NQO1 knockdown by small-interfering RNA induced degradation of both WT p53 and the p53 R273H mutant. The results indicate that curcumin induces p53 degradation and inhibits p53-induced apoptosis by an NQO1-dependent pathway.
Asher G., Tsvetkov P., Kahana C. & Shaul Y.
(2005)
GENES & DEVELOPMENT.
19,
3,
p. 316-321
Protein degradation is an essential and highly regulated process. The proteasomal degradation of the tumor suppressors p53 and p73 is regulated by both polyubiquitination and by an ubiquitin-independent process. Here, we show that this ubiquitin-independent process is mediated by the 20S proteasomes and is regulated by NQO1. NQO1 physically interacts with p53 and p73 in an NADH-dependent manner and protects them from 20S proteasomal degradation. Remarkably, the vast majority of NQO1 in cells is found in physical association with the 20S proteasomes, suggesting that NQO1 functions as a gatekeeper of the 20S proteasomes. We further show that this pathway plays a role in p53 accumulation in response to ionizing radiation. Our findings provide the first evidence for in vivo degradation of p53 and p73 by the 20S proteasomes and its regulation by NQO1 and NADH level.
Asher G., Lotem J., Tsvetkov P., Reiss V., Sachs L. & Shaul Y.
(2003)
Proceedings of the National Academy of Sciences of the United States of America.
100,
25,
p. 15065-15070
Proteasomal degradation of p53 is mediated by two alternative pathways that are either dependent or independent of both Mdm2 and ubiquitin. The ubiquitin-independent pathway is regulated by NAD(P)H: quinone oxidoreductase 1 (NQO1) that stabilizes p53. The NQO1 inhibitor dicoumarol induces ubiquitin-independent p53 degradation. We now show that, like dicoumarol, several other coumarin and flavone inhibitors of NQO1 activity, which compete with NAD(P)H for binding to NQO1, induced ubiquitin- independent p53 degradation and inhibited wild-type p53-mediated apoptosis. Although wild-type p53 and several p53 mutants were sensitive to dicoumarol-induced degradation, the most frequent "hot-spot" p53 mutants in human cancer, R175H, R248H, and R273H, were resistant to dicoumarol-induced degradation, but remained sensitive to Mdm2-ubiquitin-mediated degradation. The two alternative pathways for p53 degradation thus have different p53 structural requirements. Further mutational analysis showed that arginines at positions 175 and 248 were essential for dicoumarol-induced p53 degradation. NQO1 bound to wild-type p53 and dicoumarol, which induced a conformational change in NQO1, inhibited this binding. Compared with wild-type p53, the hot-spot p53 mutants showed increased binding to NQO1, which can explain their resistance to dicoumarol-induced degradation. NQO1 thus has an important role in stabilizing hot-spot p53 mutant proteins in human cancer.